Capitolo 2
Sull’Octopus vulgaris
2.1
Introduzione
Il soggetto particolare scelto come centro dello studio è l’Octopus vulgaris (figura 2.1), il polpo comune, una specie che, per le sue caratteristiche morfologiche, neuro-motorie e comportamentali affascina entrambi i settori di ricerca (biologico ed ingegneristico) e può fornire un chiaro esempio di come queste due discipline possono crescere in parallelo integrandosi in modo complementare.
L’Octopus mostra capacità di apprendimento e di memoria insolite per un invertebrato, e per questo è stato inteso come il paradigma dell’embodied intelligence: l’ipotesi è che l’esibizione di comportamenti intelligenti da parte di un animale che è un mollusco, sia attribuibile al fatto di essere dotato di una particolare struttura fisica, unica per le sue caratteristiche neuro-motorie e sensoriali, con cui è in grado di avere un’interazione eccellente con l’ambiente.
I tentacoli del polpo sono, per le loro particolari caratteristiche, tra quelle strutture che ad oggi catturano un elevata curiosità sia da un punto di vista biologico che robotico.
Proprio per il settore della robotica è infatti di estremo interesse la realizzazione di robot flessibili che permettono applicazioni in ambienti delicati e non strutturati [20] [21] [22]. Grande argomento di studio è anche la scelta di quale tecnica poter adottare per il loro
controllo, in m stire l’elevato
nu
colo, unita all
della morfologia e biomeccanica tentacolo, sulla base delle co
e temperati, sui fondali rocciosi lungo le coste, a non più di 100 metri di profondità. È un abile predatore solitario, che
redilige soprattutto crostacei e molluschi, con durata di vita non superiore ai due anni.
eccezione per e.
figura 2.2
o: i 2/3 dei neuroni sono infatti presenti a livello periferico, cioè nei nervi che corrono lungo i tentacoli
odo da minimizzare il numero di variabili necessarie per poter ge mero di gradi di libertà.
Per questo l’estrema destrezza che si apprezza osservando i movimenti del tenta
a conoscenza di avere alle spalle un controllo neurale piuttosto elementare, spingono la ricerca attuale verso una più approfondita conoscenza della meccanica e neurofisiologia dell’Octopus.
Detto questo, si comprende come sia necessario affrontare prima di tutto uno studio dell’anatomia generale e
noscenze biologiche attuali, per poter avere un’idea delle incredibili potenzialità che questa specie può regalare sia in termini meccanici che di controllo, e partire dalle basi biologiche per la realizzazione di sistemi artificiali di paragonabile efficienza.
2.2
Cenni di anatomia generale dell’Octopus vulgaris
L’Octopus vulgaris (phylum Mollusca, classe Cephalopoda, ordine Octopoda, famiglia Octopodidae), il polpo comune, vive soprattutto nei mari caldi
p
È caratterizzato dal non avere alcun tipo di struttura scheletrica interna, fatta il becco corneo e per un residuo rudimentale di conchiglia di dimensioni minim
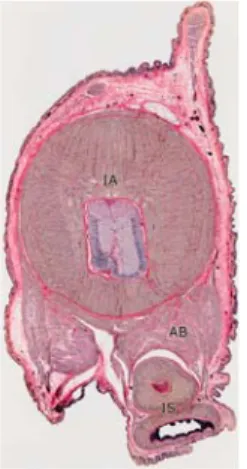
Il corpo sacciforme, di forma rotondeggiante e piuttosto voluminosa, contiene tutti gli organi vitali: il sistema nervoso centrale, 3 cuori (di cui uno pompa il sangue per il sistema vascolare e gli altri due cuori “brachiali” che pompano il sangue verso le branchie per l’ossigenazione), un rene, un sacco per l’inchiostro e uno per il veleno e lo stomaco (
).
anche due organi che gli permettono di orientarsi e di coordinare i movimenti del corpo con gli occhi: un organo analogo al nostro labirinto.
Figura 2. 2 Due rappresentazioni schematiche dell’anatomia interna dell'Octopus vulgaris.
Dalla testa emerge il sifone, che usa per resp propulsione, facendo uscire getti di acqua.
Ai lati della testa sono presenti della luce, ma non distingue i colori
Alla base inferiore della testa si trova la bocca, do
Sempre dalla base della testa, ai lati della b ca, partono le otto braccia, di lunghezza circa
uguale terno,
con cui il polpo è in grado di captare stimoli tattili e chimici.
irare, espellere gli escrementi e muoversi per gli occhi, con cui percepisce i livelli e la polarizzazione
: vede cioè in bianco e nero.
tata di un becco molto affilato. oc
e rigenerabili. Ciascun tentacolo presenta due file di ventose disposte dal lato in
Figura 2. 3 Immagine di un esemplare di Octopus vulgaris da cui si posson
ciascun tentacolo. o le due file di ventose presenti in
I tentacoli, per la caratteristica mancanza di strutture scheletriche interne, sono estremamente flessibili, virtualmente dotati di un infinito numero di gradi di libertà e, grazie anche alla presenza delle ventose, permettono al polpo di raccogliere oggetti di svariata forma e dimensione, di catturare le prede, di muoversi sul fondale marino e di arrampicarsi su scogli o pareti, oltre che di esercitare un’elevata forza di presa.
Ciascun tentacolo è controllato in modo indipendente: il sistema nervoso centrale invia dei comandi ad alto livello e quindi ciascun cordone nervoso periferico controlla il tentacolo nel dettaglio.
2.3
Morfologia del tentacolo
polpo è formato esclusivamente da tessuto muscolare, nervoso, epiteliale e connettivo, ed è il tessuto muscolare stesso che, oltre ad avere la classica funzione di generare le
ento dell’equilibrio e della postura, si
zza a riposo a seguito della contrazione dei fasci antagonisti, e deve servire da elemento di sostegno per i carichi di compressione/trazione, di flessione e di torsione esercitati dalle forze muscolari stesse.
cipali tipi di sistemi scheletrici: il primo è di tipo rigido, ed è quello presente all’interno dei vertebrati, all’esterno degli artropodi e in molti ech
no proprietà differenti, che li r
sa str
lavori manca, però, una descrizione veramente Il tentacolo del
forze necessarie al movimento attraverso la contrazione delle fibre, deve provvedere al supporto dei carichi e alla trasmissione del moto per l’intero arto.
La totale mancanza di strutture scheletriche di supporto è la prima caratteristica che rende estremamente affascinante lo studio anatomico e neuro-meccanico del tentacolo del polpo.
In genere, per l’esecuzione di movimenti, il mantenim
considera necessaria la presenza di una struttura scheletrica. Lo scheletro deve permettere la trasmissione delle forze generate dalla contrazione dei muscoli, deve assicurare il loro ritorno alla lunghe
Nel regno animale sono presenti due prin
inodermi; il secondo è invece di tipo idrostatico, tipicamente formato da tessuto connettivo con fibre di rinforzo, che racchiude al suo interno una cavità contenente fluido, e si trova ad esempio nelle specie vermiformi.
Come sarà ripetuto più volte in seguito, i muscoli del polpo han
endono organi strutturalmente unici per la loro capacità di essere sia elementi contrattili e utili al movimento, sia strutture rigide di supporto e trasmissione del movimento. La stes
uttura particolare è stata ritrovata nelle braccia e nei tentacoli di seppie e calamari, nella proboscide degli elefanti e nella lingua dei vertebrati, ed è stata definita struttura muscolare idrostatica (muscular hydrostat) [23].
In letteratura si possono trovare numerosi studi che presentano la struttura generale e l’architettura dei muscoli e dei tessuti che costituiscono il tentacolo del polpo [24], [25], [26], [27], [28], [29], [30]. In questi
pa
articolare attenzione alla traiettoria delle fibre e alla dis
e): è la superficie del tentacolo dove sono attaccate le ve
Sezione trasversale: si riferisce ai piani perpendicolari all’asse longitudinale; ♦
one centrale rispetto al tentacolo.
a intrinseca del braccio (IA), ri intrinseci di ventose e braccio [25], [31].
rticolareggiata, soprattutto per la mancanza di tecniche di indagine appropriate al tessuto in esame.
Un lavoro di recente pubblicazione [31] mette in luce nuovi dettagli per ciò che riguarda la disposizione delle fibre dei muscoli hydrostat ed il loro legame, e quindi la loro interazione, con il tessuto connettivo, con p
posizione tridimensionale.
Prima di illustrare nel dettaglio la morfologia del tentacolo è necessario chiarire alcuni termini e definizioni che saranno in seguito utilizzati:
♦ Asse longitudinale: è l’asse più lungo, che va dalla base verso la punta (tip) del tentacolo, passando per il centro;
♦ Superficie orale (oral surfac
ntose: le otto superfici orali vanno a congiungersi e racchiudersi tra loro attorno alla bocca quando l’octopus nuota in propulsione;
♦ Superficie aborale (aboral surface): è la superficie diametralmente opposta a quella orale, cioè a quella dove non sono presenti le ventose;
♦
Sezione frontale: si riferisce ai piani paralleli all’asse longitudinale e alla superficie orale;
♦ Sezione sagittale: si riferisce ai piani paralleli all’asse longitudinale, perpendicolari alla superficie orale e in posizi
Da una prima analisi visiva di una sezione trasversale del tentacolo (figura 2.4) la muscolatura interna può essere classificata in muscolatur
muscolatura intrinseca delle ventose (IS) e muscolatura acetabulo-brachiale (AB), che connette i fasci dei gruppi muscola
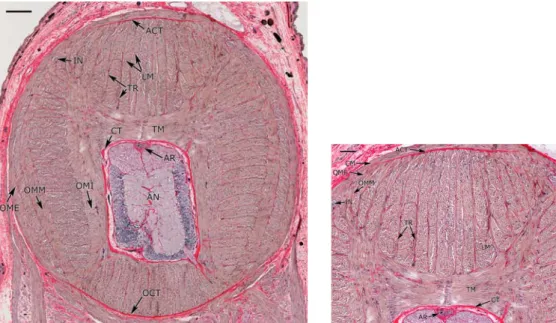
Figura 2. 4 Sezione trasversale del tentacolo in cui si può vedere la muscolatura intrinseca del braccio (IA),
La m
ristica strutturale tipica degli org
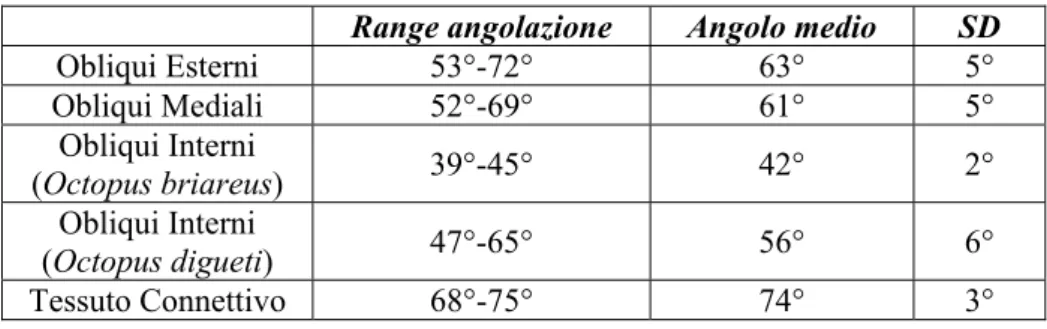
I muscoli longitudinali sono quelli costituiti dalle fibre disposte in parallelo all’asse, e hanno i fasci che corrono dalla base fino alla punta del tentacoli; i muscoli trasversali hanno i fasci disposti in direzione perpendicolare all’asse e si estendono dal centro verso la periferia; i muscoli obliqui hanno le fibre inclinate rispetto all’asse longitudinale e sono disposti per tutta la lunghezza dell’arto avvolgendolo ad elica: sono a loro volta suddivisi in obliqui esterni, obliqui mediali ed obliqui interni a seconda della loro posizione, più o meno periferica (figura 2.5).
uscolatura intrinseca del braccio è costituita da un denso impacchettamento di fibre disposte tridimensionalmente e arrangiate in fasci. È una caratte
ani con muscoli hydrostat.
Facendo quindi una prima e più generale descrizione si può suddividere le fibre muscolari in tre tipi in base al loro orientamento: parallele all’asse longitudinale, perpendicolari all’asse longitudinale e oblique, sempre rispetto all’asse longitudinale.
Figur
udinalmente l’intero arto fino alla punta. Esso comprende dei corpi cellulari disposti lateralmente e nel lato orale, e numerosi assoni di diversa dimensione disposti lungo la parte dorsale. Lungo la superficie aborale, e per tutta la lunghezza del tentacolo, si estende un’arteria con una parete piuttosto spessa. Quattro cordoni nervosi intramuscolari attraversano longitudinalmente il tentacolo e si trovano verso la periferia, nella parte più compatta della zona muscolare.
a 2. 5 Illustrazione tridimensionale di un tentacolo. Si possono vedere le diverse fibre muscolari e del
tessuto connettivo: AN, assone nervoso; AR, arteria; CM, strato dei muscoli circolari; CT, tessuto connettivo;
DCT, tessuto connettivo dermale; EP, epidermide; IN, nervo intramuscolare; LM, fibre dei muscoli
longitudinale; OME, strato dei muscoli obliqui esterni; OMI, strato dei muscoli obliqui interni; OMM, strato dei muscoli obliqui mediali; SU, ventosa (“sucker”); TM, fibre dei muscoli trasversali; TR, trabeculae; V, vena.
Al centro della sezione trasversale è ben visibile l’assone nervoso, che percorre longit
Attorno all’assone nervoso è presente uno strato spesso di tessuto connettivo (figure 2.6 e 2.8). Da studi istologici [31] le fibre che costituiscono detto strato hanno presentato la stessa reazione di colorazione tipica delle fibre di collagene. Attraverso un indagine con micrografia digitale, visto in sezione frontale o parasagittale, appare costituito da fasci di fibre intrecciate con orientamento preferenziale in direzione longitudinale e circolare. In questo strato si trovano i punti di insersione per molti fasci delle fibre dei muscoli trasversali.
Esternamente all’area occupata dalla muscolatura intrinseca del braccio sono presenti altri due spessi strati di tessuto connettivo sui lati aborale e orale, che servono come struttura di origine ed inserzione di gran parte della muscolatura intrinseca del braccio (figure 2.6 e 2.8).
sono strettamente parallele tra loro: l’orientamento è destrogiro per un set e levogiro per l’a
le fibre in un tentacolo più allungato.
ci e crostacei: co
e delle loro funzioni.
I muscoli trasversali formano una massa che circonda il cordone nervoso, nel cuore del tentacolo. Come accennato in precedenza le fibre di questo tipo sono orientate su piani Anche le fibre di questo strato hanno dimostrato di avere una reazione alla colorazione simile a quella del collagene.
In sezione frontale si osserva che queste sono raggruppate in due set vettoriali, nei quali ltro.
L’angolo che queste fibre formano con l’asse longitudinale è stato misurato pari a 68°-75° (figura 2.7). Variazioni nella misura dell’angolo sono dovute al fatto che, nonostante l’animale fosse stato anestetizzato prima dell’asportazione del tentacolo, lo stato di allungamento o accorciamento pre-morte è difficile da stabilire, e quindi si può avere un aumento di angolo in un tentacolo in uno stato di maggiore contrazione e, viceversa, un minore angolo tra
Lateralmente rispetto alla muscolatura intrinseca si trova, per ciascun lato, uno strato più sottile di tessuto connettivo, sempre utile come origine ed inserzione delle fibre muscolari, in particolare di quelle dei muscoli trasversi che si estendono attraverso il braccio.
Nel tessuto connettivo e nel tessuto epiteliale più esterno sono inclusi fasci muscolari sparsi, cromatofori, leucofori e cellule riflettenti. I cromatofori ed i leucofori sono pigmenti prodotti dal derma, molto importanti in natura per diverse specie di pes
ntengono granuli di diversi pigmenti, che in base alla loro attivazione o disattivazione portano a cambiamenti nel colore della pelle. Sono perciò le cellule responsabili delle capacità mimetiche dell’Octopus.
Tornando alla muscolatura, si fa di seguito una descrizione più dettagliata dei tre diversi raggruppamenti di fibre
pe
ci con direzione parallela al piano sagittale si estendono dalla zona orale a quella ab
ale.
rpendicolari all’asse longitudinale, in fasci approssimativamente ortogonali tra loro: una direzione parallela al piano sagittale ed una parallela al piano frontale.
I fas
orale: le fibre in questa disposizione hanno origine ed inserzione nello strato di tessuto connettivo più spesso, quello con le fibre incrociate e disposte ad elica destrorsa e sinistrorsa, e sono disposte a strati laminari (strutture definite “trabeculae” [31], vedi figura 2.6), attraversati da fasci di fibre dei muscoli longitudinali.
Alcune delle fibre trasversali che non attraversano completamente la struttura si vanno ad inserire nel tessuto connettivo che circonda il nervo centr
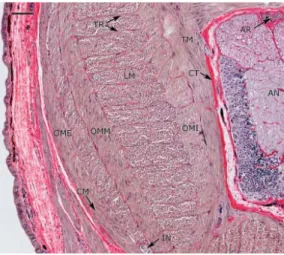
Figura 2. 6 Sezione trasversale del tentacolo (a sinistra, scale bar: 200 µm) e dettaglio della sezione stessa (a
destra, scale bar: 100 µm). Si possono vedere le componenti principali della muscolatura intrinseca del braccio:
AN, cordone nervoso assiale; ACT, strato di tessuto connettivo aborale a fibre incrociate; AR, arteria; CT,
tessuto connettivo; IN, nervo intramuscolare; LM, fibre dei muscoli longitudinali;
connettivo orale a fibre incrociate; OME, strato dei muscoli obliqui esterni; OMI, strato dei muscoli obliqui OCT, strato di tessuto inte
strato di questi muscoli ed i fasci dei lon
no invece nello strato di tessuto connettivo che circonda il nervo, senza attraversare tutta la sezione.
rni; OMM, strato dei muscoli obliqui mediali; TM, fibre dei muscoli trasversali; TR, trabeculae.
Le fibre parallele al piano frontale attraversano da un lato all’altro il tentacolo ed hanno l’origine e l’inserzione nello strato sottile di tessuto connettivo che circonda lo strato di muscoli obliqui esterni, e si estendono attraversando lo
gitudinali laterali passando sul lato inferiore (orale) e soprattutto su quello superiore (aborale) del nervo centrale (figura 2.6). Altri fasci si inserisco
Anche questi fasci formano delle trabecole laminari parallele e attraversate da fasci di muscoli longitudinali, e formano altre trabecole a fascicoli, inserendosi tra gli strati di muscoli obliqui.
Le fibre dei muscoli longitudinali si estendono parallele all’asse centrale del nervo lungo tutto il braccio e raggruppate in fasci (figura 2.7). I fasci circondano la massa centrale dei muscoli trasversali da tutti i lati (orale, aborale e laterali), con un area cross-sezionale relativa maggiore nel lato aborale rispetto a quello orale ed a quelli laterali.
La zona perimetrale esterna del tentacolo è quindi essenzialmente occupata da fasci longitudinali, separati a tratti dalle trabecole costituite dalle fibre dei muscoli trasversali e descritte in precedenza.
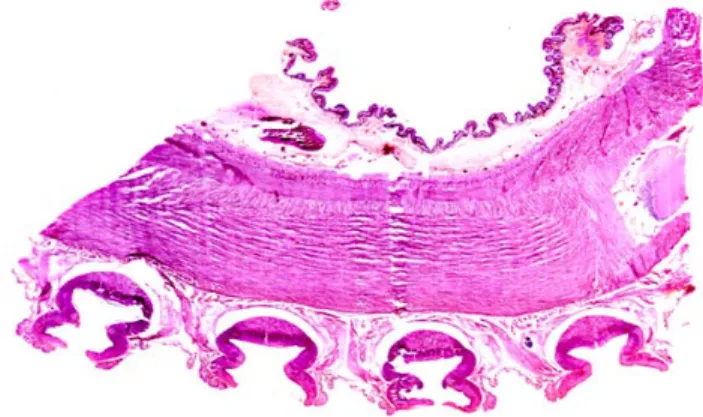
Figura 2. 7 Sezione sagittale in cui sono visibili, al centro, le fibre dei muscoli longitudinali. Nella parte
superiore si vede il tessuto connettivo della parte aborale; sotto sono invece visibili le ventose.
Uno strato di fibre longitudinali è anche presente tra i muscoli obliqui mediali e quelli esterni, anche questo è intervallato dalle trabecole dei trasversali, e presenta una forma a falce se
da una coppia di strati di fibre che si estendono da un lato e dall’altro rispetto al pia
osservato su piano trasversale (figura 2.6).
I muscoli obliqui sono suddivisi in tre set: obliqui esterni, mediali ed interni; ciascun set è costituito
no di divisione sagittale (figura 2.8).
I fasci dei muscoli obliqui esterni sono disposti nella parte più superficiale, ed avvolgono lateralmente la massa muscolare intrinseca.
Figura 2. 8 Particolare della parte laterale della sezione trasversale del tentacolo. Si vede nel dettaglio la
disposizione delle fibre dei muscoli e del tessuto connettivo. AN, cordone nervoso assiale; AR, arteria; CT, tes
obliqu
i a forma di falce descritti sopra.
Per ciascuna coppia di muscolatura obliqua (esterna, mediale ed interna) le fibre sono disposte con orientamento opposto (figura 2.10): se da un lato le fibre seguono un avvolgimento levogiro, dal lato opposto (rispetto al piano di simmetria sagittale) seguono un avvolgimento destrogiro. Inoltre, l’orientamento alternato si osserva anche tra le diverse coppie di ciascun lato: ad esempio, se da un lato i muscoli obliqui esterni sono destrogiri, dallo stesso lato si trovano i mediali levogiri e gli interni nuovamente destrogiri.
Gli obliqui esterni e mediali hanno origine ed inserzione sui lati orale ed aborale, dallo strato di tessuto connettivo spesso e con fibre incrociate, ed hanno le fibre disposte con un’angolazione simile a quella delle fibre dello stesso tessuto connettivo da cui originano.
to essere minore rispetto a quello degli altri fasci muscolari e delle fibre di tessuto connettivo.
suto connettivo; IN, nervo intramuscolare; LM, fibre dei muscoli longitudinali; OME, strato dei muscoli i esterni; OMI, strato dei muscoli obliqui interni; OMM, strato dei muscoli obliqui mediali; TM, fibre dei muscoli trasversali; TR, trabeculae.
Gli strati dei muscoli obliqui mediali si trovano in una parte un po’ più interna, sempre disposti ai due lati del corpo centrale del braccio, e sono separati dalla coppia di obliqui esterni dal gruppo di fasci di longitudinal
Gli obliqui interni sono quelli più centrali, e sono situati ai lati della massa centrale di muscolatura trasversale.
Le fibre degli obliqui interni sembra siano inserite tra le fibre dei muscoli longitudinali e trasversali, senza mostrare un chiaro punto di origine e di inserzione. L’angolo di disposizione delle fibre degli interni è risulta
Nella tabella 2.I si riportano i valori degli angoli di disposizione delle fibre dei diversi set dei muscoli obliqui. I dati di seguito riportati sono stati presi da uno studio realizzato con un
sof
e fibre del tessuto connettivo [31].
tware morfometrico attraverso cui sono stati misurati gli angoli di disposizione delle fibre dei fasci muscolari e dell
I tentacoli in esame sono stati presi da due specie diverse di polpo: l’Octopus briareus e l’Octopus digueti, che differiscono lievemente per la lunghezza dei tentacoli, più lunghi per il briareus e più corti per il digueti. I tentacoli sono stati fissati per intero e, successivamente, sono state prese 5 sezioni trasversali in punti diversi ad intervalli uguali e trattate con tecniche istologiche.
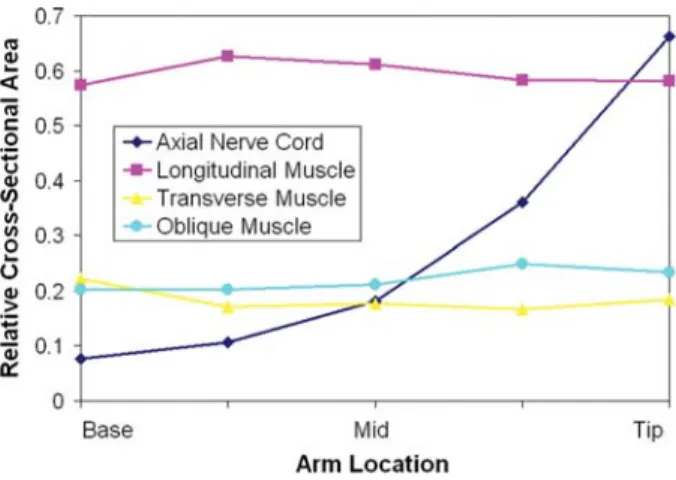
Range angolazione Angolo medio SD
Obliqui Esterni 53°-72° 63° 5° Obliqui Mediali 52°-69° 61° 5° Obliqui Interni 39°-45° 42° 2° (Octopus briareus) Obliqui Interni (Octopus digueti) 47°-65° 56° 6° Tessuto Connettivo 68°-75° 74° 3°
Tabella 2. 1 Disposizione delle fibre dei muscoli obliqui esterni, mediali ed interni e del tessuto connettivo per
l’O ten
s briareus, che ha le braccia me
orale, dove ricopre lo strato di tessuto connettivo con le fibre cross-linkate. Da questo si dirama lateralmente, ricopre i muscoli obliqui esterni e si va ad inserire nello strato di tessuto connettivo con fibre cros ora
In sezione trasversale è difficile disti strato di fibre onferenzi allo strato di fibre degli obliqui esterni perchè sono separati tra loro solo da uno strato sottile di tessuto connettivo. È invece più evidente la diff sezione paras ale, dove ota il diverso
orientamento 2.9 e 2.1
ctopus briareus e l’Octopus digueti. È definito un range di angolazione rispetto all’asse longitudinale del tacolo, ed un angolo medio con il valore corrispondente di deviatione standard.
Ciò che si nota è che tra le diverse specie di Octopus è presente un diverso grado di orientamento delle fibre degli obliqui interni: l’Octopu
diamente più lunghe rispetto alle altre specie, presenta fibre ad un angolazione inferiore rispetto all’Octopus digueti, che presenta braccia più corte e fibre a maggiore angolazione.
Molto spesso non descritti e tralasciati sono i muscoli circolari, che occupano una piccolissima parte circonferenziale (figura 2.9). Tale strato risulta leggermente più spesso nella zona ab
s-linkate della zona le.
ngere lo circ ali d
erenza in agitt si n
Figura 2. 9 Immagine della muscolatura in sezione parasagittale, leggermente di taglio rispetto all’asse
longitudinale del tentacolo. Si possono distinguere le fibre dei muscoli trasversali (TM) intrecciate ai longitudinali (LM) a livello dei muscoli obliqui mediali (OMM).
Figura 2. 10 Sezione parasagittale. Nella figura A, si vede lo strato sottile dei muscoli circolari (CM) che
circonda lo strato degli obliqui esterni (OME); accanto a questi si vede il dei longitudinali (LM) che è separato dai trasversali (TM) da uno strato di muscoli obliqui mediali (OMM): da notare la direzione opposta
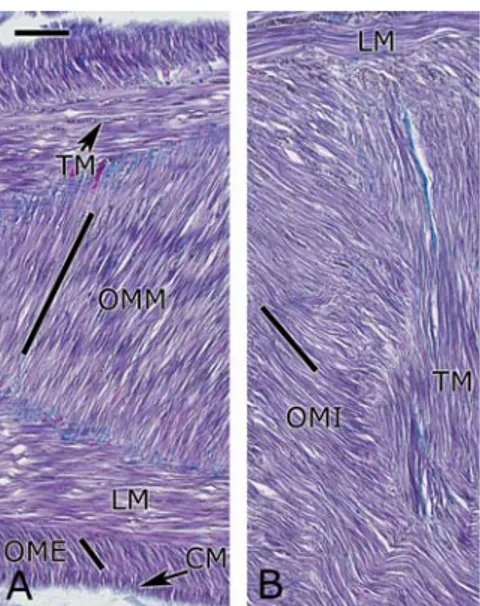
Sempre da esami istologici, sono state analizzate diverse sezioni trasversali del tentacolo su tutta la sua lunghezza. Dall’osservazione di queste è emerso che, nonostante l’area della sezione presa più vicino alla punta si riduca di 144 volte rispetto a quella presa vicino alla base, l’area relativa in sezione (proportional cross-section area, PCSA) occupata dai diversi tipi di muscoli è praticamente costante per tutta la lunghezza del braccio (figura 2.11).
fascio
tra le fibre oblique esterne e mediali. Nella figura B la sezione è stata presa a maggiore profondità: si vedono i muscoli trasversali (TM), i longitudinali (LM) e gli obliqui interni (OMI): da notare che la direzione dei fasci degli obliqui interni è la stessa dei fasci esterni della figura A.
Figura 2. 11 Grafico che mette in relazione l’area relativa cross-sezione (PCSA) occupata dai diversi fasci
muscolari lungo la lunghezza del tentacolo del polpo.
il 56 % dell’area totale, i m
nervoso, espressa in relazione all’area occupata
2.4
Biomeccanica del tentacolo
L’osservazione del tentacolo del polpo mette in prima luce la sua estrema flessibilità nell’esecuzione dei movimenti.
La struttura, essendo priva di elementi rigidi, è dotata virtualmente di un numero infinito di gradi di libertà. Quello che è stato poi osservato è che, nella pratica reale, il polpo esegue dei movimenti stereotipati e spesso riconducibili e assimilabili ai movimenti del braccio umano [33], [34], [35], [36], [37]. Resta però di fatto la potenzialità intrinseca di movimento di cui i tentacoli sono dotati.
La biomeccanica delle strutture composte da muscoli hydrostat è stata sempre più studiata negli ultimi anni [38], [39], [40].
La caratteristica fondamentale dell’elemento hydrostat è quella di mantenere un volume co
Da notare, invece, è il netto aumento di area relativa occupata dall’assone nervoso, misurata in proporzione rispetto all’area relativa occupata dalla muscolatura intrinseca.
Considerando come area totale quella occupata dalla muscolatura intrinseca, ed escludendo la porzione occupata dal nervo, i muscoli longitudinali occupano circa
uscoli obliqui il 21% e quelli trasversali il 17%. L’area relativa occupata dal tessuto dalla muscolatura intrinseca, aumenta dal 7% a livello della base, fino a circa il 59% alla punta.
Questa proprietà è legata al fatto che la struttura ha di per sé un elevato modulo di bulk (K) [31
[24]: ], che è il coefficiente che mette in relazione uno stress idrostatico, come una pressione (p), applicato ad un elemento, con la variazione di volume causata dalla pressione stessa
⎟ ⎠ ⎞ ⎜ ⎝ ⎛ = V dV K p
Si può considerare il modulo di bulk un analogo del modulo elastico E (modulo di Young) con la differenza che E mette in relazione uno stress di tipo tensile (σ) ed una deformazione ε derivante uni-dimensionale: ⎟ ⎠ ⎞ ⎜ ⎝ ⎛ = = L dL E Eε σ
Con lo stesso approccio si definisce anche il modulo di taglio G che lega shear stress e shear strain: ⎟ ⎞ ⎜ ⎛ = θ τ G d ⎠ ⎝ θ
in questo caso lo stress è applicato di taglio e la deformazione è angolare e misurata in radianti.
Le deformazioni lungo assi di riferimento differenti sono messe in relazione attraverso il modulo di Poisson: x x ε ε z y ε ε ν =− =−
ntalmente e che vale da 0 per i liquidi a 0.5 per i materiali più
ma cambiano facilmente forma (Tabella 2.II). che viene determinato sperime
solidi; quei materiali con ν → 0.5 sono quelli che hanno un’alta resistenza alle variazioni di volume,
Materiale Modulo di Poisson
Gomma ∼ 0.50 Argilla satura 0.40 - 0.50 Magnesio 0.35 Titanio 0.34 Leghe di Alluminio 0.33 Argilla 0 0 - 0.45 .3 Acciaio 0.27 - 0.31 Ghisa 0.21 - 0.26 Sabbia 0.20 - 0.45 Cemento 0.2 Vetro 0.18 - 0.30
Attraverso il rapporto di Poisson è possibile mettere in relazione i moduli precedentemente definiti: ) 1 ( 2 +ν = E G ) 2 1 ( 3 − ν = E K ed è quindi possibile scrivere:
K G G E 3 1 3 + =
Da cui si vede che il modulo elastico, E, è maggiormente legato e dipendente dalla res
costante.
La costanza di volume, dov tto che i senzialmente composti da liquido incomprimibile alla pr fisiologica [23 ì che la variazione lungo una dimensione porti ad una variazione opposta e di zione nell’altra. È questo il principio base fondamentale per le considerazioni fatte a seguire e che va sempre tenuto presente nello svolgimento di an nti le str ostat.
I muscoli hydrostat sono form bre e fasci ar diversi orientamenti, e perciò il controllo per il mantenimento del volume costante deve avvenire sulle 3 dimensioni.
Il tentacolo, attivando in maniera selettiva specifici gruppi di fibre muscolari, è in grado di
generare una larga varietà di i e compless igliore
comprensione e m movi
) e torsione.
Se si considera una struttura cilindrica con volume costante, l’allungamento sarà generato da una diminuzione dell’area della sezione trasversale. È inoltre importante sottolineare che per un elemento con un elevato rapporto lunghezza/spessore iniziale è sufficiente una piccola
dim ere un ampio aumento della lunghezza. (figura
2.12).
istenza alle azioni di taglio, piuttosto che alle variazioni volumetriche.
Il tessuto muscolare e connettivo del tentacolo possono, quindi, coerentemente godere di notevoli capacità elastiche mantenendo proprio il volume
uta al fa muscoli sono es
essione ], fa s
compensa alisi riguarda utture hydr ati da fi rangiati in
divers i movimenti. Ai fini di una m odellizzazione, questi movimenti possono essere ricondotti a cinque menti base: allungamento/accorciamento, flessione laterale (bending), irrigidimento (stiffening
Figura 2. 12 Grafico che rappresenta la relazione tra diametro e lunghezza in un cilindro di volume cost
e da cui si può vedere come una piccola diminuzione di diametro permetta un notevole aumento di lungheante zza.
-100% [22], [40]. La curiosità è che in letteratura non esistono dati qu
Alcune strutture che presentano muscoli hydrostat sono state studiate per misurare le loro performance di allungamento (Tabella 2.III). Attraverso l’applicazione di marker e l’utilizzo di videocamere, per la registrazione delle deformazioni longitudinali, si sono definite capacità di allungamento del 70
antitativi sulle performance del polpo.
Tabella 2. II Performance di allungamento misurate per diverse specie che presentano strutture
muscolari hydrostat.
Il movimento che genera l’allungamento nel tentacolo del polpo è essenzialmente e a dim nuzione della sezione trasversale e quindi un aumento complessivo della lunghezza del bra
imputabile all’azione di contrazione dei fasci dei muscoli trasversali che, com precedentemente osservato sono disposti diametralmente, e che contraendosi portano ad un
i
ccio. Come appena detto, basta una piccola contrazione dei fasci trasversali per avere un allungamento di notevole entità.
Il movimento di accorciamento è invece dovuto all’azione dei muscoli longitudinali, che con una contrazione portano ad una diminuzione della lunghezza e ad un conseguente
aumento della sezione trasversale. Detto questo appare evidente considerare i muscoli trasversali e longitudinali tra loro antagonisti di movimento (figura 2.13).
Figura 2. 13 Sezione trasversale del tentacolo. L'immagine ripresa è la stessa presente in figura 1.6; sono state
evidenziate le aree occupate dai fasci dei muscoli longitudinali, e le rette d’azione dei fasci tasversali.
Il bending è la flessione laterale del tentacolo (figura 2.14) e può avvenire su tutta la lunghezza del tentacolo ed in infinite direzioni. Il movimento di bending, in un cilindro di volume costante, può essere generato attraverso la contrazione dei fasci longitudinali da un lato
te e di avere quindi una flessione laterale (figura 2.14 A).
e un’azione di resistenza all’accorciamento dall’altro. La contrazione dei soli longitudinali porterebbe infatti all’accorciamento complessivo della struttura, con una deformazione della sezione sagittale (figura 2.14 B). La co-contrazione delle fibre trasversali o circolari del lato opposto permette invece di mantenere il diametro costan
Figura 2. 14 Diagramma illustrativo per il movimento di bending del ntacolo, o in altra struttura con
ing; nel cas tale forza resistente non è esercitata, e la struttura ha una variazione di diametro, senza il bending.
te
muscolo hydrostat. La diminuzione della lunghezza da un lato della struttura è generata dalla contrazione dei fasci dei muscoli longitudinali di quel lato. Nel caso A, il diametro è mantenuto costante dalla co-contrazione di fasci muscolari opposti, che generano una forza resistente per i longitudinali permettendo il bend
La capacità del tentacolo del polpo, così come le altre strutture hydrostat, di eseguire il bending su più piani è imputabile al fatto che i muscoli longitudinali sono posizionati nella parte periferica della sezione trasversale e lungo tutta la circonferenza (figura 2.13).
Il bending è un movimento che può essere localizzato o distribuito e che quindi implica un livello di controllo neuro-muscolare a sua volta localizzato, sia per i muscoli longitudinali che per gli antagonisti trasversali o circolari. E variando i pattern di attivazione è possibile avere il bending in punti diversi e in diverse direzioni: questa capacità dona alla struttura un’estrema destrezza e flessibilità, che oltre a dare al tentacolo la possibilità di eseguire un’infinità di movimenti complessi, permette ad esempio di svolgere tutti quei rapidi e complicati movimenti che coinvolgono la nostra lingua mentre parliamo. Il fatto che dagli esami istologici (§ 2.3) sia risultata una maggiore area di occupazione nella zona aborale fa presumere che il movimento di bending da tale zona sia quello che permette di esercitare una maggiore forza.
La torsione (twisting) del tentacolo del polpo avviene attraverso una contrazione dei fasci dei muscoli obliqui (figura 2.15) ed un’azione passiva di supporto del tessuto connettivo con le fibre incrociate.
Figura 2. 15 Rappresentazione delle rette d'azione per i muscoli obliqui responsabili della torsione
del tentacolo.
Il tentacolo del polpo, come descritto nel paragrafo precedente, comprende tre set principali di muscoli obliqui, suddivisi in base alla loro posizione osservata in sezione trasversale: gli obliqui esterni, i mediali e gli interni.
Per ciascuna coppia di muscolatura obliqua (esterna, mediale ed interna) le fibre sono disposte con orientamento opposto (figura 2.10 B): se da un lato del core le fibre seguono un avvolgimento levogiro, dal lato opposto seguono un avvolgimento destrogiro. Inoltre, l’orientamento alternato si osserva anche tra le diverse coppie di ciascun lato: ad esempio, se
da un lato i muscoli obliqui esterni sono destrogiri, dallo stesso lato si trovano i mediali levogiri e gli interni nuovamente destrogiri.
La forza prodotta dalla contrazione di una fibra di un muscolo obliquo viene trasmessa alla fibra di tessuto connettivo e da questa alla fibra sull’altro lato e così via, seguendo un percorso elicoidale lungo tutto il tentacolo.
Vista la disposizione delle fibre dei muscoli obliqui, nella torsione si genera un pattern di attivazione asimmetrico: infatti per le tre coppie di muscoli il verso di avvolgimento dell’elica
d el lato
opposto.
e si contraggono: osservando il ten
F
ento torcente dei fasci di muscoli obliqui è dato dal prodotto della forza esercitata (F
i un elemento della coppia è opposto al verso di avvolgimento dell’elemento d
Per generare una forza torsionale ci deve essere una trasmissione continua lungo tutto il tentacolo, ed è il verso di avvolgimento che determina il senso della torsione. La direzione della torsione dipende dal senso in cui sono avvolte le fibre ch
tacolo dalla base verso la punta la contrazione delle fibre avvolte ad elica destrorsa genera un movimento anti-orario della punta del tentacolo rispetto alla base (figura 1.16), viceversa una contrazione dell’elica sinistrorsa causa un movimento orario.
igura 2. 16 Movimento di torsione anti-orario per contrazione delle fibre avvolte con elica destrorsa
(osservando dalla base, in basso, alla punta, in alto).
Il mom
) per la distanza dall’asse neutro (b) :
b F
MO = ×
dove la forza esercitata è pari al prodotto tra la forza massima esercitatile e la PCSA:
PCSA
F
F
=
max×
e l’asse neutro è in generale definito come l’asse attorno a cui avviene la torsione e in cui è nullo lo sforzo di taglio, coincidente quindi con l’asse longitudinale del tentacolo.
È interessante notare che i fasci dei muscoli obliqui, sia di quelli esterni che di quelli mediali, sono posizionati nella parte più periferica del tentacolo (§ 2.3), più lontani dall’asse
neutra, e questo permette di aume ente all’atto di applicazione della forza di contrazione.
Non è invece ancora chiaro quale sia il ruolo scoli obliqui interni, troppo vicini all’asse neutra per poter sviluppare una forza a quella degli altri muscoli obliqui o perché siano disposti ad ento differente.
Quando, invece, si ge ltanea delle coppie di obliqui da entrambi i lati, quindi con pattern di attivazione simmetrico, si verifi ento della stiffness to
e fibre dei muscoli obliqui, oltre ai movimenti torsionali, partecipano anche alle va
pende dall’angolo di disposizione delle fibre rispetto all’asse longitudinale attorno cui sono avvolte.
con un certo angolo α rispetto al suo asse longitudinale.
ntare il valore del momento torc dei mu
di torsione paragonabile un angolo di orientam
nera una contrazione simu
ca un aum rsionale (usata, ad esempio, per incrementare il grado di stiffness flessionale).
L
riazioni della lunghezza del tentacolo [22]. Il loro effetto per questi movimenti di
Nella figura 2.17 si vede un cilindro di volume costante attorno cui è avvolta una fibra
Figura 2. 17 Cilindro di volume costante attorno cui è avvolta una fibra con un certo angolo α rispetto al
suo asse longitudinale.
Per il dato cilindro si hanno lunghezza, diametro e volume legati delle seguenti relazioni:
L
r
V
D
r
22
sin
cos
π
π
α
D
L
=
α
=
=
da queste si ricava che il volume, in funzione del raggio è: π α αcos
2
4 quindi, per un cilindro con volume costante:
sin 3 D V = 3 4πV /sin2αcosα D=
3 4 /sin2 cos
cosα πV α α
L=
Si riporta di seguito il grafico relativo:
Figura 2. 18 Diagramma che mette in relazione la lunghezza di una fibra avvolta ad elica attorno ad un
cilindro di volume costante con l’angolo α della stessa fibra rispetto all’asse longitudinale.
Dal grafico di figura 1.18 si vede chiaramente che il minimo della curva corrisponde ad un angolo pari a 54°44’, e cioè che la fibra avvolta attorno al cilindro di volume costante ha
lun nde al massimo per angoli che tendono a 0° e
90°.
uesto significa che una contrazione delle fibre disposte ad un angolo superiore a 54°44’
po lo inferiore
porta a torsione ed accorciamento della struttura.
Altro movimento (o caratteristica) particolarmente interessante per la struttura hydrostat, e che può essere osservato spesso tra le performance del tentacolo del polpo, è lo stiffening (o la modulazione di stiffness).
Una struttura cilindrica, con volume costante, può irrigidirsi, ovvero aumentare la propria rigidezza, se le variazioni dimensionali sono limitate da un’azione di forza resistente generata o dalla contrazione muscolare, o dalla resistenza del tessuto connettivo. La possibilità di avere una stiffness controllabile, nello spazio e nel tempo, permette al tentacolo di diventare una struttura estremamente rigida e utile come supporto (figura 2.19), e permette di eseguire movimenti complessi e combinati in diverse direzioni.
ghezza minima a tale angolo; la lunghezza te Q
Figura 2. 19 Immagine rappr
par esentativa delle diverse capacità motorie del tentacolo del polpo; in pos
ollo basate sull’utilizzo di movimenti stereotipati.
Gli studi realizzati in questo settore hanno portato alla definizione di alcuni set di movimenti stereotipati.
Il primo gruppo riguarda i movimenti di reaching, in cui si ha un aumento della distanza tra le porzioni distale e prossimale del braccio. Il reach può essere eseguito in diversi modi: uno di questi, forse il più osservato e studiato, è basato sulla propagazione di un’onda di stiffening e straightening lungo il braccio, dalla base alla punta (figura 2.20).
ticolare si può apprezzare il bending verso l’alto dei i tentacoli anteriori e lo stiffening dei tentacoli teriori che servono da sostegno.
Il meccanismo per il controllo complessivo del reclutamento dei fasci risulta necessariamente diverso da quello a noi noto e, in una sua misura, potenzialmente più sofisticato: l’ipotesi, a cui si affidano diversi gruppi di studio dell’Octopus, è che siano adottate delle tecniche per la semplificazione del contr
Figura 2. 20 Un’immagine, ripresa durante gli studi di osservazione in vivo di un esemplare di Octopus
vulgaris, in cui si osserva l’esecuzione del reaching attraverso la propagazione dell’onda di stiffness, contemporaneamente nei due tentacoli anteriori.
Un altro movimento di reaching osservato è generato dal progressivo stiramento del braccio a partire da una posizione in cui è arricciato a spirale su se stesso, fino al suo co
i può accorciare ma tenendo una struttura rettilinea o si può ripiegare con bending localizzati, conservando la lunghezza di partenza, come se fosse un braccio articolato con degli pseudo giunti (movimento di fetching).
Della terza categoria, di search/grope/explore, fanno parte quei movimenti in cui non si ha una significativa variazione nella distanza tra le porzioni distali e prossimali, ma sono movimenti laterali, di bending localizzato o torsione del braccio.
Da un punto di vista biomeccanico tutti questi movimenti sono realizzabili per mezzo delle 4 deformazioni base del braccio (allungamento, accorciamento, bending e torsione) e
he
ari punti nella sua lunghezza e possono avere diversi orientam
mpleto allungamento.
La seconda categoria comprende i movimenti di pull, in cui ho la diminuzione della distanza tra i segmenti distali e prossimali. Anche in questo caso esistono diversi modi di esecuzione: si può avere un arricciamento del tentacolo dalla punta alla base, s
n
attraverso l’utilizzo della capacità di modificare i valori di alcune delle caratteristic instrinseche della struttura, come la stiffness.
Le deformazioni possono essere singole o combinate, possono presentarsi isolate in un punto del braccio o ripetute in v
enti e direzioni (ad esempio il bending e la torsione possono essere eseguite in senso orario o in senso antiorario); contemporaneamente, la stiffness può essere variabile in tensione, compressione, flessione e torsione.