Premessa
La ricerca dell’origine della bellezza costituisce una materia importante della speculazione filosofica fin dai suoi esordi. L’estetica platonica enfatizza l’esistenza della bellezza come una presenza eterna, indipendente dal soggetto che la percepisce, sebbene sia concesso un ruolo alla partecipazione dell’individuo nell’esperienza estetica. E’ tuttavia con la pubblicazione dell’opera di Kant che l’attenzione viene spinta verso il soggetto, alla ricerca di quali presupposti diano validità e uniformità ai giudizi estetici.
La ricerca dei processi che sottostanno alla percezione della bellezza ha impegnato la psicofisiologia del diciannovesimo secolo e, più recentemente, la neuroestetica (Zeki & Lamb, 1994; Vartanian & Goel, 2003; Kawabata & Zeki, 2004), che si propone di stabilire i fondamenti biologici e neurobiologici dell’esperienza estetica. Questo studio può essere affrontato con due approcci differenti e complementari: da un lato si possono ricercare le aree cerebrali e i meccanismi fisiologici coinvolti nella percezione della bellezza, dall’altro indagare quali caratteristiche di uno stimolo siano in grado di suscitare tale percezione.
Una visione evoluzionistica della materia, inoltre, spinge a ricercare quali forze selettive abbiano condotto all’evoluzione dell’esperienza estetica. In quest’ottica lo studio della percezione della bellezza nei volti e nei corpi degli esseri umani offre il vantaggio di poter essere inquadrato all’interno della teoria della selezione sessuale, uno dei cui scopi è l’identificazione dei meccanismi evolutivi che hanno portato alle preferenze per determinate caratteristiche nei potenziali partner riproduttivi e sessuali. Negli ultimi decenni numerosi studi hanno indagato il comportamento sessuale umano con un approccio evoluzionistico; molte ricerche sono state dedicate all’identificazione delle componenti biologiche associate all’attrazione, soprattutto per quanto riguarda la bellezza dei volti, che costituiscono per la percezione umana degli stimoli di particolare significato.
Selezione sessuale
Secondo Darwin, la selezione sessuale deriva da differenze nel successo riproduttivo causate da competizione per i partner. L’idea della selezione sessuale è sorta dal tentativo di Darwin di risolvere uno dei maggiori problemi della sua teoria dell’evoluzione per selezione naturale, riguardante i caratteri vistosi posseduti dai maschi di alcune specie che sembrerebbero ridurre la sopravvivenza e dovrebbero quindi essere osteggiati dalla selezione naturale. Per risolvere questo paradosso Darwin ha proposto una distinzione tra selezione naturale e selezione sessuale, basata quest’ultima non su una lotta per la sopravvivenza, ma sulla competizione per il possesso di partner riproduttivi (Darwin, 1859). Il termine “competizione” si riferisce a tutte le situazioni in cui l’uso di una risorsa (in questo caso, un partner riproduttivo) da parte di un individuo rende la risorsa più difficile da ottenere per altri individui. La competizione per i partner caratterizza quindi tutte le forme di selezione sessuale, inclusa la scelta del partner, in cui gli individui di un sesso competono per essere scelti da individui del sesso opposto (Andersson, 1994).
Un carattere è sottoposto a selezione sessuale quando la sua espressione è legata a differenze nel successo riproduttivo causate da competizione per i partner. Queste differenze possono derivare da molte forme di competizione diretta o indiretta per l’accoppiamento, che includono scontri diretti, forme di resistenza, “gare di velocità”, scelta del partner e competizione spermatica.
Un meccanismo di selezione sessuale che ha ricevuto particolare attenzione è la scelta del partner da parte di un sesso. Questo meccanismo non si riferisce ovviamente a un processo cognitivo, ma all’insieme di caratteri e comportamenti che rendono più probabile l’accoppiamento con alcuni partner potenziali piuttosto che con altri (Kirkpatrick & Ryan, 1991). Nella maggior parte delle specie animali la selezione sessuale è più pronunciata nel sesso maschile e la scelta del partner ricade sul sesso femminile; tuttavia il tipo di sistema nuziale, le differenze nell’investimento parentale, il potenziale riproduttivo e la sex-ratio possono influenzare la forza della selezione sessuale nei due sessi, e sotto alcune condizioni la competizione e la scelta del partner possono essere osservati in entrambi i sessi (Andersson, 1994).
Un numero elevato di osservazioni ed esperimenti sia in natura che in laboratorio hanno fornito evidenza che le femmine spesso scelgono il proprio partner ripoduttivo e che tale scelta favorisce particolari caratteri maschili (Andersson, 1982; 1994). Tuttavia i meccanismi evolutivi che hanno condotto all’evoluzione delle preferenze femminili sono ampiamente dibattuti (Andersson & Simmons, 2006) e la difficoltà è aumentata dalla possibilità che diversi meccanismi intervengano insieme.
Sono stati proposti diversi meccanismi per spiegare l’evoluzione delle preferenze nella scelta del partner, cui sono stati dedicati modelli matematici teorici e ricerche empiriche:
Effetti fenotipici diretti. La preferenza femminile per un ornamento maschile può evolversi come risultato di benefici fenotipici diretti che la femmina (o in generale il sesso che compie la scelta) ottiene dal partner riproduttivo. I benefici possono essere di molteplice natura: risorse materiali che il partner è in grado di offrire alla femmina o alla prole, protezione, risparmio di tempo nella ricerca di un partner, ridotta trasmissione di malattie o parassiti.
Tendenze sensoriali (o sfruttamento sensoriale). Un tratto maschile può essere favorito perchè si trova a stimolare una tendenza già esistente nel sistema sensoriale femminile, evolutasi per selezione naturale per rispondere ad altre esigenze, ad esempio alimentari o antipredatorie. Sono ora disponibili numerose evidenze dell’esistenza di tendenze sensoriali in un crescente numero di specie, dove analisi filogenetiche hanno mostrato che la preferenza femminile per un carattere maschile è presente anche in altre specie imparentate in cui manca il carattere maschile (Endler & Basolo, 1998; Ryan, 1998). Processo di Fisher (o processo runaway o dei figli sexy). Questa idea, avanzata da Fisher nel 1930, propone che le elaborate esibizioni maschili possano essere selezionate semplicemente perchè rendono i maschi attraenti per le femmine. L’ipotesi prevede che l’accoppiamento genetico tra gli alleli che codificano per la preferenza femminile e gli alleli che codificano per il tratto maschile possa portare, sotto alcune condizioni, ad una coevoluzione auto-rinforzante tra carattere e preferenza. Se questo accoppiamento genetico si diffonde abbastanza nella popolazione, le femmine traggono un vantaggio in termine di fitness dalla scelta di un maschio che esprime un determinato carattere perchè i figli, ereditando il carattere, saranno preferiti da altre femmine.
Meccanismi indicatori (o dei buoni geni o dell’handicap). Se i caratteri sottoposti a selezione sessuale sono espressi in proporzione alla condizione del possessore, un carattere ben sviluppato dovrebbe indicare una buona condizione e vitalità. Nella misura in cui lo stato di salute e la vitalità sono ereditabili, una preferenza femminile per un tratto ben sviluppato aumenta la probabilità che la progenie erediti la qualità genetica del padre. Questa teoria si basa sull’assunto, proposto da Zahavi, che i tratti sottoposti a selezione sessuali siano costosi, e costosi in modo differenziale per individui di diversa qualità genetica (Andersson, 1994). Una difficoltà che pone questa teoria dipende dalla definizione del concetto di qualità genetica e dai criteri utilizzati per misurarla (Hunt et al., 2004), soprattutto perchè si ritiene che la condizione di un individuo sia influenzata da gran parte del suo genoma (Tomkins et al., 2004). Un altro aspetto dibattuto riguarda il mantenimento della variabilità genetica, o “paradosso del lek”, presupposto necessario perchè ci siano vantaggi genetici ereditabili. Una possible soluzione a questo problema è stata offerta dall’idea che la selezione non operi in modo uniforme e che il fenotipo ottimale vari nello spazio e nel tempo, ad esempio attraverso meccanismi coevolutivi tra ospiti e parassiti. L’ipotesi dei parassiti, avanzata da Hamilton e Zuk (1982), postula una correlazione tra i geni per la resistenza ai parassiti e l’espressione del carattere sottoposto a selezione sessuale: le femmine
con una preferenza per il carattere tenderanno ad accoppiarsi con maschi più resistenti ai parassiti. La correlazione tra la resistenza ai parassiti e l’espressione del tratto potrebbe spiegarsi con il costo che caratterizza quest’ultima in termini di immunocompetenza. E’ stata dimostrata in alcune specie l’esistenza di un compromesso tra l’investimento in segnali sessuali maschili e funzione immunitaria (Blount et al., 2003; Faivre et al., 2003).
Meccanismi di compatibilità genetica. Piuttosto che scegliere un partner con “buoni geni”, una femmina potrebbe mostrare una preferenza per un partner con un patrimonio genetico compatibile con il proprio. Questa teoria è stata invocata come possibile spiegazione della poliandria in alcune situazioni, soprattutto sotto l’azione della competizione spermatica: se esiste una variabilità maschile nella compatibilità genetica con una femmina, e se ci sono dei meccanismi che rendono più probabile la fecondazione delle uova da parte di sperma più compatibile, o se le femmine possono investire più risorse in embrioni fecondati da maschi più compatibili, allora la compatibilità genetica può spiegare gli accoppiamenti femminili con più maschi (Tregenza & Wedell, 2000). Numerosi esempi di scelta del partner basata su compatibilità genetica sono stati osservati nel caso dei geni MHC, un cluster di geni principalmente coinvolti nella regolazione della risposta immunitaria.
I meccanismi che sono stati proposti per spiegare l’evoluzione delle preferenze femminili sono mutualmente compatibili e possono verificarsi insieme. Con il passare delle generazioni le preferenze femminili sono in grado di modificare la morfologia della specie ed in alcune condizioni a dare origine a speciazione (Higashi et al., 1999; Haesler & Seehausen, 2005).
La percezione dei volti
Numerose ricerche indicano che i volti rappresentano per la percezione umana degli oggetti molto particolari, cui sono dedicati circuiti cerebrali specializzati, coinvolti nell’analisi selettiva delle proprietà visive delle facce.
L’esistenza nel cervello umano di sistemi neurali specializzati per la percezione dei volti fu suggerita inizialmente dall’osservazione di pazienti con danni cerebrali focali che mostravano una selettiva incapacità di riconoscere volti familiari, mentre erano in grado di riconoscere correttamente altri oggetti. Questa sindrome è chiamata prosopagnosia ed è associata a lesioni molto spesso bilaterali che coinvolgono la corteccia occipitale ventrale e la superficie interna dei lobi temporali (vedi Haxby et al., 2000 per una review).
La dissociazione tra meccanismi neurali per la percezione dei volti e degli oggetti è stata supportata dagli studi condotti sul cosiddetto “effetto di inversione” sia in soggetti normali che in pazienti con deficit selettivi nel riconoscimento di volti o di oggetti. Le persone possono imparare rapidamente a riconoscere immagini di volti mai visti prima, ma incontrano notevoli difficoltà nell’imparare a riconoscere immagini di volti quando queste sono presentate capovolte. Al contrario, il riconoscimento di immagini di altri oggetti non è influenzato dall’inversione delle immagini (Yin, 1970).
Yin e altri ricercatori hanno quindi ipotizzato l’esistenza di circuiti specializzati nell’analisi delle caratteristiche che permettono di distinguere un volto da un altro. L’inversione dell’immagine di un volto, modificando la configurazione globale delle caratteristiche visive, impedisce a questi circuiti specializzati di funzionare al meglio. Nei pazienti affetti da prosopagnosia, d’altra parte, l’inversione dei volti non causa una performance peggiore nel compito di riconoscimento, suggerendo che la percezione di volti invertiti in questi pazienti possa essere mediata dai loro meccanismi intatti di percezione di oggetti. Diversi esperimenti condotti su di un paziente con agnosia per gli oggetti hanno mostrato che la percezione dei volti era normale ma il riconoscimento di volti invertiti mostrava un deficit severo (Moscovitch et al., 1997), suggerendo che i meccanismi per il riconoscimento di volti non possano processare con successo i volti invertiti.
Un’ulteriore evidenza proviene da studi su primati non umani: nei macachi sono stati identificati dei neuroni nel solco temporale superiore e nella corteccia infero-temporale che rispondono selettivamente ai volti. Questi risultati hanno suggerito che un’area dedicata specificamente alla percezione dei volti potesse esistere in regioni omologhe del cervello umano (vedi Haxby et al., 2000 per una review). Lo sviluppo delle tecniche di brain imaging funzionale ha permesso di studiare in modo non invasivo le regioni del cervello umano coinvolte nelle percezione dei volti con una precisione anatomica maggiore di quanto sia possibile in pazienti con lesioni cerebrali (Kawinsher et al., 1996).
Numerose ricerche indicano che la percezione di volti evoca l’attivazione di una regione del giro fusiforme laterale, una circonvoluzione cerebrale localizzata nel lobo temporale inferiore. L’attivazione è generalmente bilaterale, ma più consistente nell’emisfero destro (Clark et al., 1996; Haxby et al., 1999). Quest’area corticale mostra un’attivazione maggiore durante test di percezione di volti piuttosto che durante test che coinvolgono la percezione di stimoli di controllo o di altri oggetti.
Altre regioni coinvolte nella percezione dei volti sono state identificate nel giro latero-inferiore occipitale e nel solco postero-superiore temporale.
L’utilizzo di fMRI in studi sull’effetto di inversione ha mostrato che i volti invertiti continuano a evocare l’attivazione dei sistemi neurali deputati alla percezione dei volti, ma coinvolgono anche regioni della corteccia extrastriata che rispondono maggiormente ad altri oggetti. L’inversione di immagini di oggetti, invece, non causa un simile aumento di attivazione in regioni che rispondono ai volti. La partecipazione
di regioni generalmente dedicate alla percezione di altri oggetti nella percezione di volti invertiti può spiegare perchè questi stimoli sembrino essere processati più come non-volti e perchè i pazienti affetti da prosopagnosia non mostrino un peggioramento nei compiti di percezione dei volti invertiti (Haxby, 1999).
E’ stato ipotizzato che il sistema neurale dedicato alla percezione dei volti sia suddiviso in moduli funzionalmente differenti in grado di processare diverse caratteristiche degli stimoli; in particolare sono stati distinti gli aspetti invarianti di un volto, che caratterizzano l’identità di una persona, e gli aspetti mutevoli, quali l’espressione emotiva, la direzione dello sguardo, i movimenti legati al linguaggio (Haxby et al., 2000; Hoffman e Haxby, 2000; Phillips et al., 1997).
La percezione della bellezza
La bellezza nei volti umani è stata a lungo considerata all’interno della categoria generale della teoria estetica e solo recentemente nel dominio della biologia e delle neuroscienze.
Alcune ricerche indicano che i bambini fin dai primi mesi di età mostrano una preferenza visiva per volti giudicati attraenti dagli adulti rispetto a volti non attraenti, e che tale preferenza è diretta verso volti attraenti piuttosto che verso volti simmetrici quando queste due variabili sono considerate in modo indipendente (Samuels et al., 1994; Slater et al., 1998). Questa evidenza rende improbabile che la percezione della bellezza sia un fattore puramente legato all’apprendimento di una particolare convenzione culturale.
Inoltre è stato osservato un generale consenso nei giudizi di attrazione attribuiti ai volti, sia all’interno degli stessi gruppi sociali che tra differenti culture e società (Perrett et al., 1994; Perrett et al., 1998; vedi Thornhill & Gangestad, 1999 per una review).
Alla luce di queste considerazioni, negli ultimi anni alcuni studi di brain imaging hanno ricercato quali aree cerebrali siano coinvolte nella percezione della bellezza.
Un primo studio (Nakamura et al., 1998) ha utilizzato la tomografia a emissione di positroni per confrontare l’attivazione cerebrale di soggetti maschili durante l’esecuzione di compiti differenti di fronte all’esposizione di una serie di immagini di volti femminili: in un test ai soggetti è stato chiesto di ricoscere il colore dello sfondo, in un altro test di giudicare l’emozione del volto e infine di attribuire un giudizio di attrazione. Si è osservata un’attivazione maggiore nelle regioni frontali della corteccia sinistra durante l’attribuzione del giudizio di attrazione rispetto alle altre due condizioni. All’interno di queste
aree la regione frontale anteriore ha mostrato un’attivazione correlata con il giudizio di volti poco attraenti, mentre la corteccia orbito-frontale si è attivata durante il giudizio di volti attraenti.
In una ricerca successiva (Aharon et al., 2001) sono stati analizzati tramite fMRI soggetti maschili sottoposti a visione passiva di fotografie di volti maschili e femminili che erano stati precedentemente giudicati come molto attraenti o normali. La visione di volti attraenti ha causato un’attivazione selettiva della corteccia orbitofrontale destra, dell’amigdala estesa sublenticulare destra e del tegmento ventrale bilaterale, mentre la visione dei soli volti femminili attraenti ha attivato in modo specifico il nucleus accumbens, una regione cerebrale coinvolta nel sistema della ricompensa. Questo risultato ha suggerito che possa esistere una differenza tra una percezione puramente estetica e una percezione di uno stimolo coinvolto in un meccanismo di ricompensa, come può essere un volto di un individuo del sesso opposto.
Uno studio ha osservato un’attivazione dello striato ventrale correlata positivamente con la bellezza solo quando lo sguardo dei volti giudicati è diretto verso i soggetti; la correlazione diventa negativa quando lo sguardo è diretto altrove, causando una diminuzione dell’attivazione dello striato ventrale all’aumentare della bellezza (Kampe et al., 2001).
Un altro gruppo di ricerca ha riscontrato che la visione di volti attraenti produce attivazione nella corteccia orbitofrontale mediale, in un contesto in cui i soggetti non sono chiamati esplicitamente a giudicare la bellezza dei volti (O’Doherty et al., 2003). Inoltre il grado di attivazione in questa regione è risultato modulato dalla misura in cui i soggetti hanno percepito la presenza di un’espressione positiva nel volto osservato: i volti attraenti che sono stati percepiti come sorridenti hanno causato una risposta maggiore nella corteccia orbitofrontale rispetto ai volti con espressione neutrale. Altre regioni hanno mostrato attivazione durante la visione di volti attraenti: la corteccia prefrontale mediale e la corteccia del cingolo posteriore. E’ stata inoltre osservata una risposta differente nella corteccia orbitofrontale mediale e laterale: mentre la corteccia orbitofrontale mediale ha mostrato un’attivazione maggiore alla visione di volti attraenti, la regione laterale ha risposto maggiormente a volti poco attraenti. Secondo gli autori questo effetto è consistente con precedenti evidenze di una dissociazione tra risposte nella corteccia orbitofrontale mediale e laterale a diversi tipi di rinforzi: ricompense e punizioni (O’Doherty, 2001).
I correlati geometrici dell’attrazione nei volti
Ben prima che fossero identificate le regioni cerebrali coinvolte nella percezione della bellezza, numerosi ricercatori si sono impegnati nella ricerca dei tratti dei volti correlati con l’attrazione.
Una caratteristica dei volti che è stata associata con la bellezza è la somiglianza al volto medio.
Questo dato emerse in modo inaspettato da uno studio di Sir Francis Galton alla fine del XIX secolo; cercando di estrarre le caratteristiche tipiche di una serie di volti (uno dei suoi propositi era di ottenere il prototipo del criminale) egli creò con un sistema di sovrapposizione fotografica dei volti compositi che erano la fusione di più volti reali. I volti ottenuti con questa procedura si mostrarono stranamente reali e più belli delle fotografie individuali che componevano le immagini (vedi Etcoff, 1994).
Il risultato è stato confermato da studi più recenti che si sono avvalsi di una versione più moderna della tecnica per la realizzazione di volti medi. I volti creati con programmi di morphing mediante la fusione di una serie di volti reali sono stati giudicati più attraenti con l’aumentare del numero di immagini utilizzate per creare gli stimoli, conducendo i ricercatori all’affermazione che la bellezza dei volti risiedesse nella somiglianza alla media (Langlois et al., 1990).
Questa preferenza può essere spiegata considerando la selezione stabilizzante, che agendo contro gli estremi della popolazione favorirebbe i possessori di caratteristiche medie, che risulterebbero essere dei partner riproduttivi ideali; la tendenza a trovare tali caratteristiche attraenti si troverebbe quindi ad essere altrettanto favorita.
Altri ricercatori, meno inclini a pensare che i meccanismi psicologici umani siano stati plasmati dall’evoluzione, hanno fornito spiegazioni diverse per la preferenza per i volti medi, interpretandola invece come un sotto-prodotto di una più generale caratteristica del sistema percettivo che conduce alla preferenza per stimoli di varia natura, ad esempio cani, uccelli o orologi, che presentano caratteristiche medie (Halberstadt & Rhodes, 2000).
Un volto medio tuttavia, per quanto attraente, potrebbe non essere il più attraente. E’ quanto è risultato da uno studio (Perrett et al., 1994) che ha indagato le preferenze per tre tipi di stimoli realizzati con un programma di morphing: un volto medio ottenuto dalla fusione di un certo numero di fotografie, un volto ottenuto solo dalla fusione dei volti giudicati più attraenti e un volto ottenuto accentuando le differenze tra i due precedenti stimoli. Il viso ottenuto dai volti più attraenti è stato preferito rispetto al viso medio, ma accentuando le differenze si è ottenuto un viso giudicato ancora più attraente, quanto meno per i volti femminili.
Questo studio ha indicato che esistono delle caratteristiche geometriche associate in modo specifico con la bellezza.
Una caratteristica estesamente analizzata come possibile indicatore della bellezza dei volti è la simmetria bilaterale. L’asimmetria fluttuante è costituita dalla deviazione dalla simmetria per tratti che sono simmetrici a livello della popolazione ed è considerata un indice di instabilità durante lo sviluppo. Per un organismo è infatti particolarmente costoso creare due metà perfettamente identiche, resistendo a stress ambientali e genetici, e in questo modo la simmetria potrebbe risultare un segnale onesto della qualità dell’individuo. Assumendo che la stabilità durante lo sviluppo sia almeno in parte ereditabile, una preferenza per la simmetria nella scelta del partner potrebbe essere favorita dalla selezione.
Gli studi che hanno testato questa ipotesi, tuttavia, hanno mostrato risultati contradditori, riportando correlazioni positive (Grammer & Thornhill, 1994; Perrett et al., 1999; Scheib et al., 1999; Penton-Voak et al, 2001) o negative (Swaddle & Cuthill,1995, Zaidel et al., 2005) tra la bellezza e la simmetria dei volti. La differenza tra questi risultati può essere imputabile ai diversi metodi utilizzati per creare gli stimoli (vedi Thorhill & Gangestad, 1999 per una discussione) o per misurare l’asimmetria nei volti. Un’altra fonte di variabilità riguarda ovviamente il grado di asimmetria presente nel campione di soggetti utilizzati: mentre risulta infatti intuitivo che una forte asimmetria (che significa però anche una forte lontananza dal volto medio) sia negativamente correlata con la bellezza, è più difficile stabilire con certezza se le leggere fluttuazioni nei tratti dei volti abbiano un valore altrettanto negativo nel giudizio di attrazione. Inoltre uno studio interessante (Scheib et al, 1999) ha suggerito che la simmetria possa essere associata alla percezione della bellezza senza essere necessariamente un fattore che la influenza: è stata infatti osservata una correlazione tra il giudizio di attrazione e la simmetria misurata nei volti, ma tale relazione non è scomparsa quando ai soggetti sono state presentate immagini della metà destra o sinistra dei volti. Questo risultato suggerisce che altre caratteristiche del volto, correlate con la simmetria, possano influenzare il giudizio di attrazione.
Alcuni ricercatori hanno fornito una spiegazione differente per la preferenza per stimoli simmetrici, sostenendo che la simmetria sia più facilmente percepita nel sistema visivo. Secondo questa ipotesi la preferenza per volti simmetrici non sarebbe che un sottoprodotto di una generale tendenza a trovare più piacevoli oggetti con simmetria bilaterale. Uno studio ha tuttavia indicato che la preferenza per volti resi simmetrici rispetto alla versione naturale e asimmetrica degli stessi volti viene meno quando gli stimoli vengono presentati invertiti (Little & Jones, 2003), suggerendo che la simmetria dei volti sia processata da un meccanismo specifico a causa del forte significato adattativo di questa capacità. Quest’ipotesi sembra essere confermata da un altro studio (Rhodes et al., 2005) che ha osservato che la performance nel riconoscimento dell’asimmetria nei volti è significativamente peggiorata quando i volti vengono presentati invertiti, con i negativi delle immagini o con un filtraggio delle immagini.
Un altro fattore considerato importante nell’attribuzione del giudizio di attraenza è il dimorfismo sessuale. Nella specie umana i due sessi differiscono per una serie di caratteri sessuali secondari sia anatomici che fisiologici: la dimensione del corpo, la distribuzione del grasso corporeo e il metabolismo lipidico, la quantità e la distribuzione di pelo corporeo, il colore della pelle, la tonalità di voce e alcune caratteristiche facciali.
In molte specie animali i caratteri sessuali secondari, la cui espressione si trova spesso sotto controllo ormonale, sono oggetto di selezione sessuale (Andersson, 1994). Il possesso di tali caratteri potrebbe essere indice della buona condizione dell’individuo, e se l’espressione del tratto varia con la condizione fenotipica e genotipica del possessore, una preferenza per quel tratto si troverebbe ad essere favorita. Negli esseri umani alcune caratteristiche estrogeno-dipendenti del corpo femminile sono considerate segnali di salute e di successo riproduttivo e sono giudicate attraenti (Barber, 1995).
Nell’uomo l’espressione di molti caratteri sessuali secondari è testosterone-dipendente; il grado di espressione di questi caratteri è proposto da alcuni ricercatori come un onesto indicatore di qualità dell’individuo a causa di una putativa azione immuno-soppressiva del testosterone (Bouman et al., 2005).
Anche il volto mostra un chiaro dimorfismo sessuale: il volto maschile mostra mandibola e mento più larghi orizzontalmente, sopracciglia più basse, maggiore prominenza nell’arcata sopraccigliare e nel ponte nasale, che fanno apparire gli occhi più infossati e più piccoli rispetto agli occhi delle donne. Il volto femminile invece si presenta con naso relativamente più piccolo e concavo, sopracciglia più alte e arcuate, labbra più carnose e la parte inferiore del viso più corta e stretta.
Per testare l’effetto del dimorfismo sessuale sulla percezione della bellezza uno studio ha realizzato con delle tecniche di morphing un volto medio maschile e un volto medio femminile, modificabili lungo un continuum in modo da accentuare o diminuire le caratteristiche dimorfiche (Perrett et al., 1998). E’ stato poi chiesto a dei soggetti di entrambi i sessi di modificare i volti fino ad ottenere il volto giudicato più attraente. Per quanto riguarda il volto femminile, soggetti di entrambi i sessi hanno scelto una versione femminilizzata del volto medio, confermando l’ipotesi che l’accentuazione dei caratteri sessuali secondari sia correlata con la bellezza. Il giudizio sul volto maschile ha invece fornito un risultato meno prevedibile, con una preferenza per un volto maschile femminilizzato rispetto al volto medio. Per interpretare queste preferenze, gli autori dello studio hanno successivamente chiesto a dei soggetti di giudicare il volto medio, il volto femminilizzato e il volto mascolinizzato secondo una serie di tratti della personalità: l’accentuazione delle caratteristiche maschili è risultata associata ad un aumento nella percezione della dominanza, della mascolinità e dell’età, ma ad una diminuzione nella percezione di alcune caratteristiche positive quali l’affidabilità e l’onestà. E’ stato quindi suggerito che la preferenza per un volto maschile femminilizzato possa essere mediata sia dalla preferenza per delle caratteristiche
infantili, che dall’evitamento di caratteristiche associate a tratti psicologici negativi. Queste preferenze indicherebbero una pressione selettiva sull’evoluzione della forma del viso che limita un dimorfismo sessuale pronunciato nei visi maschili ma non femminili.
Uno studio successivo (Penton-Voak et al., 1999) ha indicato che il giudizio delle donne su volti maschili potrebbe essere influenzato dai cambiamenti ormonali associati al ciclo mestruale. Anche in questa ricerca è emersa una generale preferenza per volti maschili femminilizzati. Tuttavia durante il periodo fertile del ciclo mestruale le donne intervistate hanno mostrato una preferenza per un minore grado di femminilizzazione rispetto al periodo non fertile, soprattutto quando è stato loro richiesto di valutare il volto come possibile partner in una relazione a breve termine; nel giudicare invece il volto come partner a lungo termine il momento del ciclo mestruale non ha mostrato un effetto significativo sulla preferenza. Secondo gli autori questo risultato potrebbe essere interpretato come una strategia di scelta del partner mista, in grado di ottimizzare i vantaggi della preferenza secondo le condizioni in cui si trova la donna: i tratti più maschili, indice di buona immuno-competenza, sarebbero preferiti nella condizione in cui il rischio di concepimento è più alto.
Altri studi, tuttavia, non hanno confermato la preferenza per volti maschili femminilizzati. Un altro gruppo di ricerca ha utilizzato un metodo differente per valutare l’impatto delle caratteristiche dimorfiche sul giudizio di attraenza (Johnston et al., 2001): invece di creare un volto ipermaschile accentuando le differenze tra volto medio maschile e volto medio femminile, è stato utilizzato un programma di algoritmi genetici che permette di modificare singole caratteristiche dei volti muovendosi all’interno di uno spazio delle facce multidimensionale. E’ stato chiesto a dei soggetti di utilizzare gli algoritmi genetici per selezionare il volto percepito come più maschile e più femminile e queste immagini sono state poi utilizzate per creare un continuum di mascolinità e femminilità percepita. I volto giudicato più attraente lungo questo continuum era significativamente più mascolino rispetto al volto medio maschile. Anche questo studio ha comunque osservato uno spostamento della preferenza verso un volto più mascolino durante la fase fertile del ciclo mestruale, indicando inoltre che il giudizio di attraenza sui volti maschili è l’unica componente a variare in modo significativo durante il ciclo mestruale, mentre restano costanti i giudizi sui volti femminili e l’attribuzione di caratteristiche psicologiche ai volti maschili.
Questi studi tuttavia hanno valutato come varia il giudizio di attrazione su un volto medio al variare del dimorfismo sessuale, cosa ben diversa dal considerare i suoi effetti su di un viso reale. Alcuni studi realizzati su volti reali hanno misurato un indice di mascolinità basato su alcune caratteristiche facciali dimorfiche (ad esempio, la prominenza degli zigomi e le dimensioni della mandibola), osservando una correlazione tra l’attrazione e la mascolinità così misurata (Grammer & Thornhill, 1994; Scheib et al., 1999). Altri ricercatori (Swaddle & Reierson, 2002) hanno modificato delle immagini di volti maschili reali in modo da imitare le differenze che risultano da diversi livelli di testosterone durante la pubertà. Sono
stati realizzati in questo modo dei continuum che trasformassero i volti reali in modo da accentuare o diminuire le caratteristiche maschili; è stato poi chiesto a dei soggetti femminili di indicare l’immagine giudicata più attraente e quella percepita come più dominante: mentre la dominanza è risultata associata ad un aumento delle caratteristiche maschili, la scelta del volto più attraente si è concentrata sulle immagini più vicine al volto non modificato. Gli autori hanno interpretato questi risultati come l’effetto di una selezione stabilizzante sui livelli di testosterone presenti nella popolazione ed espressi nei volti maschili.
L’osservazione di questi studi sembra indicare che la mancanza di concordanza nei risultati derivi dalla difficoltà di trovare un indice opportuno che quantifichi il grado di dimorfismo sessuale nei volti e possa quindi essere correlato con il giudizio di attrazione.
La percezione del dimorfismo sessuale
Il riconoscimento di genere è una facoltà fondamentale per la vita sociale di un individuo. Gli esseri umani sono in grado di discriminare il sesso di un’altra persona sulla base di numerose informazioni visive riguardanti le dimensioni e la forma del corpo, la quantità e la distribuzione dei peli sul corpo, la forma del viso e una serie di artefatti culturali quali pettinatura, cosmetici e abbigliamento.
Il volto umano, che in molte società è l’unica parte del corpo sempre visibile, è in grado di fornire informazioni sufficienti per il riconoscimento del sesso: anche dopo la rimozione di tracce culturali (capelli, trucco, gioielli), una fotografia del volto porta ad una corretta discriminazione del sesso dell’individuo quasi nel 100% dei casi (Bruce et al., 1993). Per indagare quali informazioni siano utilizzate per questo compito discriminativo, diversi studi hanno investigato l’effetto della rimozione o della riduzione delle fonti di informazioni che potrebbero essere usate. E’ stato indagato il ruolo svolto dalle tracce superficiali, quali texture e colore della pelle, e dalla geometria del viso, intesa sia nei suoi elementi distinti (occhi, naso, mento...) che nella configurazione globale delle caratteristiche del volto. Per studiare l’effetto della texture sulla discriminazione del sesso sono state utilizzate immagini di volti sottoposti a scanner laser, in modo da conservare soltanto le informazioni riguardanti la forma del viso (Bruce et al., 1993; Hill et al., 1995). Il riconoscimento del sesso di questi stimoli è risultato meno accurato e dipendente dall’orientamento del volto da giudicare, con una performance peggiore per la visione frontale rispetto al profilo e ai ¾, soprattutto nella discriminazione dei volti femminili (Bruce et al., 1993). Questo dato suggerisce che le informazioni convogliate dalle caratteristiche superficiali del volto
siano importanti nel riconoscimento del sesso, soprattutto quando, come nella visione frontale, la geometria tridimensionale del viso è più difficile da valutare.
Per indagare il ruolo svolto dai singoli elementi del volto nel riconoscimento del sesso sono stati utilizzati approcci differenti: il mascheramento di singole parti del viso di fotografie (Bruce et al., 1993), la presentazione di elementi isolati del volto (Brown & Perrett, 1993) e la creazione di volti medi maschili e femminili con la sostituzione di elementi del prototipo del sesso opposto (Brown &Perrett, 1993; Yamaguchi et al., 1995). Nell’insieme questi studi indicano che le parti del volto che hanno un maggior peso nell’attribuzione del sesso sono la regione degli occhi e delle sopracciglia e la regione della mandibola.
E’ stata inoltre osservata un’influenza del sesso del volto nel compito discriminativo: i volti maschili risultano più facilmente riconoscibili dei volti femminili, soprattutto quando viene ridotta l’informazione visiva presentando i volti come scanner laser (Bruce et al., 1993) o tramite delle tecniche di filtraggio delle immagini (Cellerino, 2004). Questa differenza non sembra dovuta ad una tendenza percettiva a categorizzare un volto come maschile quando lo stimolo è ambiguo: sono stati creati stimoli non-senso ottenuti dall’alternanza di strisce di volti maschili e femminili e presentati con un livello di filtro delle immagini che rendesse invisibile la presenza delle strisce. Di fronte a questi stimoli la discriminzione del sesso è risultata non statisticamente differente dal caso, senza mostrare quindi una tendenza a categorizzare un volto ambiguo come maschile (Cellerino, 2004). Sembra pertanto che il volto maschile abbia bisogno di meno informazioni per essere riconosciuto correttamente.
Si è riscontrata inoltre una differenza nel riconoscimento di genere nei due sessi, con una migliore performance dei soggetti femminili nel riconoscere volti femminili (Cellerino, 2004). Un più efficiente riconoscimento di sesso in volti appartenenti allo stesso sesso del soggetto giudicante è stato osservato anche in uno studio che ha utilizzato volti e soggetti giapponesi (Yamaguchi et al., 1995). Inoltre, i soggetti femminili sono risultati più efficienti rispetto ai soggetti maschili nel riconoscimento di volti femminili in un test di memorizzazione dei volti (Lewin & Herlitz, 2002).
Considerando dunque che le numerose informazioni convogliate da un volto hanno un diverso peso nella percezione del dimorfismo sessuale, risulta evidente che per valutare l’influenza del dimorfismo sessuale sul giudizio di attrazione è suggeribile utilizzare non solo un indice metrico delle caratteristiche facciali tipiche dei due sessi, ma anche una variabile che tenga conto dell’aspetto percettivo di tali caratteristiche.
Morfometria geometrica
La morfometria geometrica è un metodo per l’analisi multivariata della forma (shape). Mentre il termine form si riferisce ad una combinazione di shape e size (forma e dimensione), il termine shape indica l’insieme delle informazioni geometriche che rimangono quando gli effetti della scalatura, della posizione e della rotazione sono eliminati da un oggetto. La sua formulazione teorica si è sviluppata negli ultimi decenni dalla sintesi di metodi di analisi multivariata di matrici di covarianza e metodi per la visualizzazione diretta di cambiamenti nella forma biologica (Bookstein, 1996). Questa analisi consente di preservare l’integrità della forma biologica in due o in tre dimensioni, evitando che essa si riduca ad una serie di misure lineari e angolari che non includono informazioni riguardanti le relazioni geometriche dell’intero oggetto. Questa proprietà ha reso la morfometria geometrica uno strumento molto utilizzato nelle analisi antropometriche (Ponce de Léon & Zollikofer, 2001; Hennessy & Stringer, 2002; Rosas & Bastir, 2002; Hennessy et al., 2005; Pretorius et al., 2006).
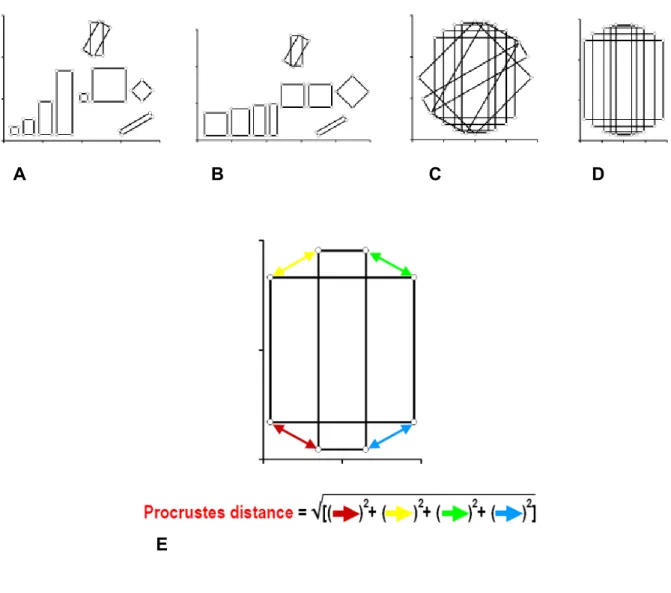
La morfometria geometrica descrive la forma come un set ordinato di punti omologhi chiamati landmark (Bookstein, 1996). Per confrontare le configurazioni di landmark delle immagini è necessaria una serie di trasformazioni matematiche che ne ottimizzano la sovrapposizione: l’ Analisi Generalizzata di Procuste trasforma le configurazioni tramite traslazione, scalatura e rotazione dei sistemi di coordinate. La traslazione porta a coincidere nello spazio i centroidi di tutte le configurazioni di coordinate, la scalatura ne normalizza la taglia, infine la rotazione ne ottimizza l’orientamento secondo una procedura di minimi quadrati. Dopo queste trasformazioni, che costituiscono la superimposizione di Procuste, le differenze di forma possono essere descritte dalle differenze in coordinate dei landmark corrispondenti negli oggetti, che diventano in questo modo disponibili per analisi multivariate (Fig. 1.1).
La distanza di Procuste, che quantifica la differenza di forma tra le configurazioni di landmark, è descritta dalla radice quadrata della somma delle distanze al quadrato tra i landmark corrispondenti dopo la superimposizione di Procuste. La forma media del campione può essere definita come la forma che ha la minore distanza di Procuste dalle forme individuali (Fig. 1.1).
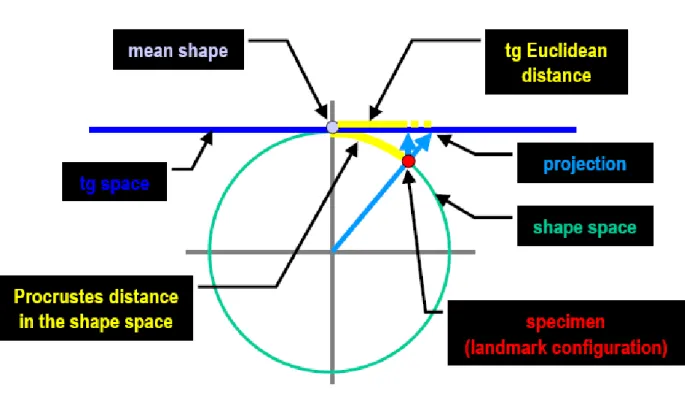
I metodi della morfometria geometrica sono basati sullo spazio multidimensionale delle forme definito da Kendall, in cui ogni forma del campione può essere considerata come un punto, o un vettore. Lo spazio di Kendall è uno spazio curvo, basato su una geometria non-euclidea; tuttavia quando la variazione della forma è relativamente piccola è possibile utilizzare uno spazio lineare tangente lo spazio di Kendall nel punto rappresentato dalla forma media, così come una mappa può approssimare in modo accurato una piccola regione della Terra (Rohlf, 1998) (Fig. 1.2).
La morfometria geometrica permette di visualizzare le differenze di forma come deformazioni di una griglia cartesiana capace di interpolare i cambiamenti nelle conformazioni di landmark minimizzando la
bending energy attraverso la funzione della thin plane spline (TPS). Un’analisi spettrale di questa funzione fornisce le sue autofunzioni, chiamate principal warp. I coefficienti di queste autofunzioni, chiamati partial warp, costituiscono una base ortonormale per lo spazio morfometrico.
I principal warp possono essere utilizzati come input per un’Analisi delle Componenti Principali (PCA), che calcola i principali assi di variazione nello spazio tangente. Le PCA possono essere visualizzate tramite thin plane spline e possono essere messe in relazione con supposte proprietà biologiche, fornendo la possibilità di visualizzare variazioni della forma in funzione di ipotetiche variabili biologiche.
A B C D
E
Fig. 1.1 In alto: Schematizzazione del metodo di superimposizione di Procuste. (A) Vengono raccolte le coordinate dai
dati grezzi. (B) Le configurazioni di landmark vengono scalate alla stessa dimensione. (C) La traslazione delle configurazioni di landmark porta a far coincidere nello spazio i centroidi. (D) La rotazione minimizza le distanze tra landmark corrispondenti. In basso: Rappresentazione della distanza di Procuste, descritta dalla radice quadrata della somma delle distanze al quadrato tra landmark corrispondenti dopo la superimposizione di Procuste.
Fig. 1.2 Rappresentazione dello spazio delle forme di Kendall, approssimato da uno spazio euclideo tangente lo
Scopo della tesi
In questo lavoro è stata applicata la morfometria geometrica per esplorare le correlazioni tra l’attrazione e la geometrica del volto maschile.
Trentatré profili maschili e 33 profili femminili sono stati utilizzati per creare uno spazio morfometrico multidimensionale all’interno del quale è stato possibile identificare una misura della somiglianza al volto medio (distanza di Procuste) ed una misura del dimorfismo sessuale geometrico.
Per eliminare l’effetto della texture sul giudizio, sono stati realizzati mediante un programma di morphing (GrkMorhp) stimoli a texture uniforme, ottenuti adattando alla geometria dei volti individuali una texture maschile media.
Le immagini dei 33 profili maschili sono stati sottoposti al giudizio di soggetti femminili, chiamati a rispondere secondo una scala di Likert a 7 possibilità (da 1=molto brutto a 7=molto attraente).
E’ stata inoltre valutata una componente, chiamata Femminilità Percepita, che fornisse un indice della percezione del dimorfismo sessuale. A questo scopo i profili maschili ed i profili femminili sono stati presentati con texture androgina in un test di riconoscimento del sesso.