4. I CARBONATI MICROBIALITICI ED
IL "PROBLEMA DOLOMITE"
La gran parte dei carbonati microbialitici e non, è composta di calcite, alto o basso magnesiaca, o di aragonite; quelli a composizione dolomitica, totale o parziale, sono a priori considerati “dolomitizzati” attraverso fasi diagenetiche più o meno precoci. Questa sorte di pregiudizio diagenetico nei confronti dei carbonati dolomitici derivava dalla effettiva quasi totale mancanza negli ambienti attuali di dolomite "primaria", cioè di dolomite di precipitazione diretta da un fluido in condizioni ambientali (bassa temperatura e bassa pressione). Tale possibilità, recentemente messa in luce proprio da studi su carbonati microbialitici, ha aperto nuove vedute sulla formazione della dolomite e sulla genesi degli ingenti volumi di carbonati dolomitici presenti nel record fossile (Burns et al., 2000).
4.1. LA PRECIPITAZIONE DELLA DOLOMITE PRIMARIA ATTRAVERSO LA MEDIAZIONE BIOLOGICA
Il nome originale del minerale, dolomie, era stato dato da N.T. Saussare, nel 1792, in onore del geologo francese Deodat Guy de Dolomieu 1750–1801, che per primo la descrisse in rocce delle Alpi Tirolesi. A più di due secoli dalla sua scoperta rimangono ancora aperti molti interrogativi riguardanti soprattutto la sua origine e la sua distribuzione attraverso il record geologico, riferiti ad una controversia nota come “il problema dolomite”.
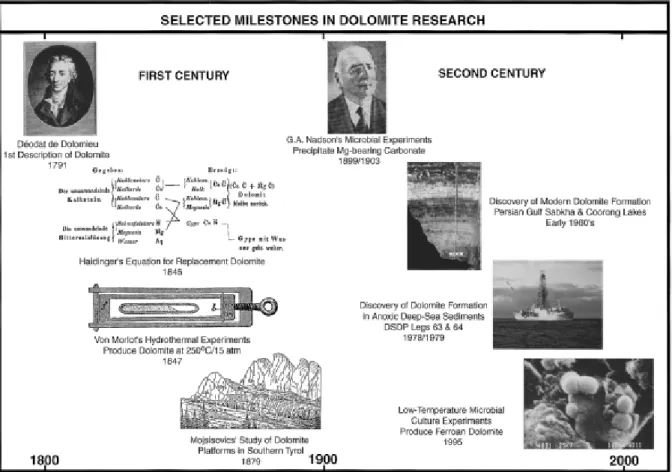
Nel primo secolo di ricerca si cercarono di spiegare i principi fisici e chimici di dolomitizzazione, basati principalmente sulla comune osservazione che esisteva una genetica associazione tra dolomite e gesso (Haidinger, 1845) (Fig. 4.1). Il secondo secolo ha visto nuove scoperte di ambienti moderni dolomitici e lo sviluppo di nuove idee che sono state formulate all’interno di vari modelli. Restavano ancora aperti, tuttavia, molti problemi riguardo sia alla distribuzione irregolare della dolomite nel record geologico sia all'incapacità di indurre la precipitazione della dolomite in laboratorio, in condizioni simili a quelle trovate negli ambienti moderni dolomitici (McKenzie 1991).
La dolomite si trova in grande abbondanza nelle rocce antiche a partire dal Precambriano e per buona parte del Fanerozoico; nel Quaternario e nell'Attuale la sua diffusione risulta invece limitata. Sebbene siano stati proposti numerosi modelli per la formazione della dolomite (Hardie, 1987), rimane ancora un enigma la spiegazione di una
così grande disuguale distribuzione. La dolomite attuale si trova in una larga varietà di ambienti: da quelli lacustri a quelli subtidali, fino ai quelli marini profondi. Molti dei modelli idrologici proposti per la formazione di dolomite in acque basse coinvolgono il pompaggio, attraverso i sedimenti, di larghi volumi di acqua di mare a salinità normale, oppure di brine ipersaline (Adams & Rhodes, 1960; Carballo et al., 1987; Vahrenkamp et al., 1991; Swart & Melim, 2000; Melim & Scholle, 2002). Altri modelli prevedono la formazione di dolomite in una zona di mescolamento di acqua marina con acqua meteorica (mixing zone) (Badiozamani, 1973; Humphery & Quinn, 1990; Humphery & Radjef, 1991; Humphrey, 2000). Questi modelli, in ogni modo, non spiegano sufficientemente come la precipitazione della dolomite superi i fattori cinetici inibitori, così come l'alta energia d'idratazione del Mg 2+, l'estrema bassa attività degli anioni CO32-, e la necessaria presenza di basse concentrazioni di solfati.
Figura 4.1. Schema dei principali passi nella ricerca sul "problema dolomite" (Burns et al., 2000).
In seguito alla scoperta di dolomite in sedimenti pleistocenici ricchi di carbonio organico, durante progetti di deep-sea drilling condotti nel Golfo della California (DSDP Leg 63, Pisciotto & Mahoney, 1981; DSDP Leg 64, Kelts & McKenzie, 1982), è stato proposto un
modello organo-genico per la formazione di dolomite in ambienti marini profondi. Questo modello propone la precipitazione di dolomite attraverso il simultaneo aumento nell'ambiente dell'alcalinità e il quasi azzeramento della concentrazione di solfati, attraverso la degradazione della materia organica operata dai batteri solforiducenti (Irwin, 1980; Kelts & McKenzie, 1982; Compton, 1988). Tale modello, comunque, non può essere invocato per spiegare l'origine delle grandi masse di dolomite delle piattaforme carbonatiche del Precambriano e del Fanerozoico.
Dolomite "primaria", o penecontemporanea, è stata scoperta in diversi ambienti lacustri e marini di acqua bassa nelle piattaforme carbonatiche delle Bahamas, del Golfo Persico, Florida, nei laghi del Coorong Region (Australia) (Deffeyes et al., 1965; Illings et al., 1965; Shinn et al., 1965; Shinn, 1968; Von der Borch & Jones, 1976; Von der Borch & Lock, 1979; Patterson & Kinsman, 1982; Carballo et al., 1987; Rosen et al., 1988; Rosen & Coshell, 1992; Wright, 2000). In questi ambiti la dolomite è associata con sedimenti ricchi di materia
organica, o con tappeti algali (microbici) sepolti che si trovano allo stesso livello, o in prossimità di quelli dove precipita la dolomite.
A questo punto, partendo dal riconoscimento che la dolomite possa formarsi in condizioni anossiche, la potenziale importanza della materia organica, nonché della riduzione dei solfati operata da microbi nella precipitazione della dolomite, è stata generalmente accettata (Garrison et al., 1984; Burns et al., 1988). Ma, dopo aver riconosciuto che la materia organica era probabilmente un fattore importante per la precipitazione della dolomite, rimaneva ancora da chiarire la sua funzione precisa (Purser et al., 1994).
Attraverso studi basati su metodi geomicrobiologici, Vasconcelos et al. (1995) documentano un concreto link tra la formazione della dolomite e i microbi. Questi autori segnalano i risultati di un esperimento di una coltura batterica in cui viene prodotta dolomite ricca in Fe, in condizioni anaerobiche ed a bassa temperatura, parzialmente simili a quelli riscontrati in Lagoa Verhmelha (Brasile), un ambiente attuale di precipitazione di dolomite. Come step successivo, Vasconcelos & McKenzie (1997) propongono un nuovo modello per la formazione della dolomite, basato sui loro studi nel laboratorio naturale di Lagoa Verhmelha (Brasile). Il loro modello, basato principalmente sul ruolo dei batteri nella precipitazione della dolomite, fornisce un sistema attualistico differente con cui valutare le formazioni antiche di dolomite presenti nel record geologico (Wright, 1997).
L'importanza dei batteri per la precipitazione della dolomite non è, tuttavia, un’idea così recente. Iniziando nel 1899, Nadson (1928) condusse esperimenti con colture batteriche nelle quali usò batteri solfato-riducenti, prelevati dalla superficie anossica dei sedimenti di un lago
salato in Russia, per produrre piccole quantità di un carbonato autigeno. Analisi chimiche qualitative di questi carbonati lo portarono a proporre l’idea che la dolomite potesse precipitare in collaborazione con batteri anaerobici. Circa mezzo secolo più tardi, Neher (1959) segnalò la precipitazione di cristalli di dolomite in un esperimento condotto con batteri anaerobici. Recentemente, Folk (1993) è stato un forte fautore dell'influenza dei batteri sulla formazione dei minerali, basandosi sull’osservazione di sferoidi nanometrici all'interno e sulle facce dei cristalli, interpretati come remains di antichi batteri.
Anche se il link tra batteri e dolomite è stato riconosciuto già moltissimi anni fa, l'importanza del fattore microbico nella formazione della dolomite è stato generalmente trascurato e non gli è stata data la legittima considerazione all'interno della voluminosa letteratura compilata sul “dolomite problem”.
Recentemente, tuttavia, sulla base dei risultati ottenuti da esperimenti in colture batteriche, e sull’osservazione diretta di minerali carbonatici precipitati all’interno di living mats, è stato proposto che la dolomite primaria possa essere considerata come un biominerale, ovvero un prodotto metabolico di processi biologici, quali la riduzione e l'ossidazione batterica del solfato (van Lith et al., 2003a; Visscher & Stolz, 2004; Wright & Wacey, 2005) (Fig. 4.2).
Figura 4.2. Modello schematico in 5 fasi, che illustra il ruolo dei batteri riducenti e di quelli solfo-ossidanti nella formazione di Mg-carbonati all'interno di un microbial mat (van Lith et al., 2003a)
Quindi, nella stessa ottica, essendo le strutture stromatolitiche l'annotazione sedimentaria dell’attività microbica fotosintetica (cianobatterica), la dolomite micritica potrebbe rappresentare una firma geobiologica (riduzione batterica del solfato) dell’attività microbica, probabilmente connessa con condizioni di alta alcalinità, ipersalinità ed anossia. Inoltre, i segnali geochimici registrati nella dolomite microbica, quali i rapporti dell'isotopo del carbonio e specifiche molecole organiche intercristalline, possono essere usate per documentare o confermare la sua origine microbica.
La dolomite micritica è presente in modo diffuso in tutto il record geologico, perfino nella roccia sedimentaria più vecchia, con un'età almeno di 3.45 Ga. Inoltre, la quantità relativa di dolomite rispetto al calcare e la relativa distribuzione riguardo al tempo geologico mostrano delle distinte variazioni. Per esempio, la dolomite era particolarmente comune nel Precambriano, mostra variazioni cicliche durante il Fanerozoico ed è rara nell’Olocene. Applicando il modello microbico della dolomite a questa record, Mckenzie & Vasconcelos (2001) associano i periodi di aumento di deposizione della dolomite con i livelli elevati di anossia globale e con un cambiamento nel metabolismo dominante nell'oceano. Sulla base di questa interpretazione microbica del record geologico della dolomite, propongono, inoltre, che la dolomite micritica possa essere considerata come un minerale fossile in cui sono registrate le condizioni ambientali verificatesi durante lo sviluppo della Terra.
4.2. LA DOLOMIA PRINCIPALE Fm – STILI DI DOLOMITIZZAZIONE
La Formazione della Dolomia Principale, o Hauptdolomit, del dominio Alpino-Appenninico rappresenta un caso di “Law of Persistence of Facies” (Ager, 1973), ovvero la diffusa presenza di determinate facies su ampie zone durante uno specifico intervallo del tempo geologico.
L’Hauptdolomit Formation è stata definita da Gümbel (1861) nelle Alpi Calcaree Settentrionali sulla base di caratteristiche morfologiche e litologiche, particolarmente evidenti nelle montagne delle Dolomiti. Facies simili, conosciute come Formazione della Dolomia Principale, sono state trovate in tutta la penisola italiana, ed anche nella Spagna meridionale, ed ancora in Grecia, Turchia, Ungheria, Dinaridi e Carpazi.
La Dolomia Principale consiste di una successione ciclica dolomitizzata, potente circa 1000 metri, caratterizzata da facies subtidali e peritidali di piattaforma interna, con faune limitate. La sua sedimentazione ha preso posto su un’ampia piattaforma pericontinentale in
condizioni semiaride, a partire dal Carnico superiore (?), attraverso tutto il Norico ed, in alcune aree, il Retico (De Zanche, 1990). Questo tipo di sedimentazione ha “omogeneizzato” la paleogeografia Ladinico-Carnica, caratterizzata dalla presenza di molte piattaforma carbonatiche e di bacini relativamente poco profondi. Durante il Retico, l’estensione delle facies di piattaforma della Dolomia Principale si è invece cospicuamente ridotta fino alla sua scomparsa nel Lias inferiore.
Gli studi sulla dolomite, concentratisi sulla Dolomia Principale, hanno apportato grandi cambiamenti verso la comprensione generale della genesi di imponenti successioni dolomitiche.
Studi comparativi sulle successioni dolomitiche del Triassico superiore delle Dolomiti del Brenta (ed aree adiacenti) e dei Monti Lattari (Appennino Meridionale) hanno mostrato che il Norico e il Retico sono caratterizzati da due differenti stili di dolomitizzazione (Iannace & Frisia, 1994). Questo sembra supportato anche da dati presenti in letteratura concernenti il Triassico superiore delle aree peri-Mediterranee (Ungheria: Haas, 1988; Balog et al., 1991 – Dinaridi: Cadet, 1975 – Ellenidi: Hofbauer, 1985 – Carpazi: Michalik, 1980).
Da questi lavori risulta che le successioni carbonatiche di shelf del Norico sono affette da un fabric cristallino fine, con preservazione delle strutture originali. Sulla base di dati di campagna, questa dolomitizzazione si sarebbe completata quasi interamente nel Norico. Al contrario, i carbonati retici sono solo parzialmente dolomitizzati, con successioni dolomitiche massive ed un fabric cristallino distruttivo. Queste facies si presentano come grandi corpi irregolari che tagliano la stratificazione. Inoltre, alcuni segnali isotopici ed osservazioni al TEM indicano una temperatura relativamente alta durante la sostituzione. La dolomite con fabric cristallino fine, che si ritrova comunemente nelle facies inter-sopratidali sottostanti, in questo intervallo di tempo è molto più rara; le relazioni di intersezione con i cementi sin-sedimentari indicano, comunque, una sua origine primaria rispetto a quella saccaroide.
Gli Autori ipotizzano, dunque, che la Dolomia Principale Norica si sia formata come il risultato di processi avviati da particolarissime condizioni geografiche e sedimentologiche che non hanno controparti moderne. La distribuzione della dolomite nei sedimenti dei bacini intrapiattaforma e la sua relazione con le facies progradanti di piattaforma, suggerisce che i fluidi dolomitizzanti si siano originati in ambienti marini evaporitici, come indicato dall’alto valore del δ18O. Il sistema idrologico potrebbe essere simile a quello del “reflusso” di Adams & Rhodes (1960) o al “Modello della sabka” di Müller et al. (1990), cioe un flusso di acque più dense e più saline dalla piattaforma interna verso i bacini. La persistenza nel tempo e nello
spazio di tali condizioni potrebbe spiegare la formazione del gigante corpo dolomitico della Dolomia Principale.
Al contrario la dolomitizzazione del Retico non sembra essere stratigraficamente costretta e può essere più facilmente spiegata dagli attuali modelli di dolomitizzazione. Per esempio, il modello di dolomitizzazione “Florida-Bahamas” potrebbe essere applicato alla dolomite con fabric cristallino fine, osservata nelle facies inter-sopratidali. I corpi dolomitizzati irregolari, affioranti come patches all’interno delle successioni Retiche, andrebbero invece considerati singolarmente, e possibilmente relativi ad una circolazione di fluidi a larga scala, che potrebbe essere stata attiva dal Retico in poi.
La differenza nello stile di dolomitizzazione, rivelata da questi studi, diventa più significativa quando è inquadrata all’interno dello scenario sedimentario della Tetide occidentale nel Trias superiore. Infatti, la totale consistenza dello stile di dolomitizzazione dei sedimenti norici su una così ampia regione implica che questo fenomeno sia stratigraficamente controllato, ovvero relativo allo specifico intervallo di tempo. Al contrario, nel Retico non tutte le successioni di piattaforma sono dolomitizzate, e la maggior parte dei corpi dolomitici tagliano la stratificazione. Inoltre, le relative facies di dolomite microcristallina primaria sono di gran lunga meno comuni. Queste differenze suggeriscono che al passaggio Norico-Retico si siano verificati importanti cambiamenti ambientali che interessarono i mari poco profondi della Tetide occidentale. Notevoli modifiche ambientali interessarono tutti i mari poco profondi della Tetide, dal Norico superiore all’Hettangiano. La transizione dalla “Persistenza delle Facies” della Dolomia Principale Norica alla più complessa distribuzione di facies Liassica, è il risultato di un improvviso aumento dell’attività tettonica, che ha preso posto al passaggio Norico-Retico e che può essere vista come il preludio dell’apertura dell’Oceano Ligure-Piemontese (Dewey et al., 1973; Laubscher & Bernoulli, 1977; Catalano & D’Argenio, 1982; Jadoul, 1985). Da un punto di vista globale, questo processo è parte della frammentazione del Pangea, verificatesi dal Carnico fino al Lias medio, che ha completamente cambiato il disegno della distribuzione oceani/continenti influenzando, di conseguenza, il clima e la paleocircolazione sull’intera Terra (Veevers, 1989).
A scala globale il limite Triassico-Giurassico è visto come un periodo di cambiamenti di primo ordine nel clima (Fischer, 1981; 1984), nella paleooceanografia (MacKenzie & Piggot, !981), nella mineralogia dei sedimenti carbonatici (Sandberg, 1983), nel contenuto atmosferico di CO2 (Cuif & Gautret, 1991), etc., Tutte queste teorie suggeriscono una
variazione eustatica di primo ordine (Vail et al., 1977) come causa principale di questi cambiamenti.
Il cambiamento di facies osservato può anche essere considerato come un’espressione della trasgressione a scala mondiale registrata nel Retico. Evidenze paleontologiche nell’Europa nord-occidentale suggeriscono che i mari trasgressivi erano caratterizzati da bassa salinità (Hallam & El Shaarawy, 1982). Una rapida trasgressione ed un input clastico caratterizzano i depositi retici delle Alpi e dell’Appennino settentrionale, mentre nell’Appennino meridionale mancano i sedimenti terrigeni. Questo suggerisce che la fonte di rifornimento dell’input clastico sia stato il Nord del supercontinente Laurasia, e che la sua causa sia imputabile all’aumento dell’apporto fluviale a seguito di un cambiamento di clima. E’ storicamente risaputo, infatti, che il Retico marca una transizione da un clima arido, tipico del periodo Norico, verso un clima più umido. L’aumento dell’apporto fluviale ha avuto anche l’effetto di abbassare la salinità nella Tetide occidentale. Mentre, i sedimenti clastici non raggiungevano il suo margine meridionale perché venivano raccolti nei bacini intrapiattaforma.
In conclusione, in un quadro di cambiamenti globali verificatesi tra il Trias superiore ed il Giurassico inferiore, il Retico è certamente il periodo più critico per questa transizione. Questo è particolarmente vero nel regno Tetideo, ma è supportato anche dalla stratigrafia classica del Triass francese e germanico, dove sedimenti marini retici, stanno direttamente sopra il Keuper evaporitico (Carnico e Norico). Inoltre, nel record stratigrafico dell’Europa nord-occidentale, il Retico ha delle caratteristiche paleontologiche e litologiche non riscontrabili nel Triassico e nel Giurassico (Wells, 1959).