Capitolo 3
Il ruolo delle specie reattive
dell’ossigeno nelle piante
3.1 Duplice ruolo delle ROS nella biologia degli organismi
Quando l’O2 è coinvolto in reazioni di trasferimento di elettroni, esso viene convertito in varie molecole estremamente reattive, denominate complessivamente specie reattive dell’ossigeno. Questo processo prevede la riduzione sequenziale dell’O2 ad anione superossido (O2-), perossido di idrogeno (H2O2) e radicale idrossile (OH-), specie tossiche che possono portare alla distruzione ossidativa della cellula. Conseguentemente, quindi, all’evoluzione degli organismi fotosintetici evolventi l’O2, le ROS sono divenute compagne inopportune della vita aerobica e, di conseguenza, la sopravvivenza di tali organismi risulta dipendente dallo sviluppo di efficienti meccanismi di detossificazione (Halliwell, 1989).
Nonostante le ROS siano state considerate per molti anni esclusivamente dei composti tossici, recenti risultati hanno dimostrato un loro coinvolgimento nella regolazione e il controllo di processi biologici quali crescita, ciclo cellulare, morte cellulare programmata (programmed cell death, PCD), risposta a stress (a)biotici e sviluppo (Neill et al., 2002). Questi studi suggeriscono un duplice ruolo delle ROS nella biologia degli organismi, sia come prodotti secondari tossici del normale metabolismo aerobio cellulare, sia come fattori chiave di regolazione di crescita,
sviluppo e difesa (Mittler et al., 2004), tanto da essere definite dei "master hormone" (Slesak et al., 2007).
Il bilancio tra produzione e metabolismo delle ROS appare, quindi, estremamente critico per il destino della cellula e lo “scoppio ossidativo” sembra essere un aspetto comune nella risposta della pianta agli agenti (a)biotici. Di conseguenza, la delucidazione dei meccanismi intercellulari che mediano la segnalazione tramite ROS è potenzialmente interessante per ogni lavoro che tende ad incrementare la tolleranza agli stress nelle specie coltivate.
Sotto il termine ROS sono raggruppate un gran numero di molecole, tra queste l’H2O2 risulta la più stabile e sembra poter viaggiare per alcune distanze molecolari. Inoltre, recenti risultati hanno dimostrato la presenza di specifiche acquaporine in piante e mammiferi in grado di permettere il trasporto dell’H2O2 attraverso le membrane, rendendo possibile il passaggio dal sito di origine al luogo di azione e/o detossificazione (Bienert et al., 2006).
3.2 Formazione delle ROS nei diversi compartimenti cellulari
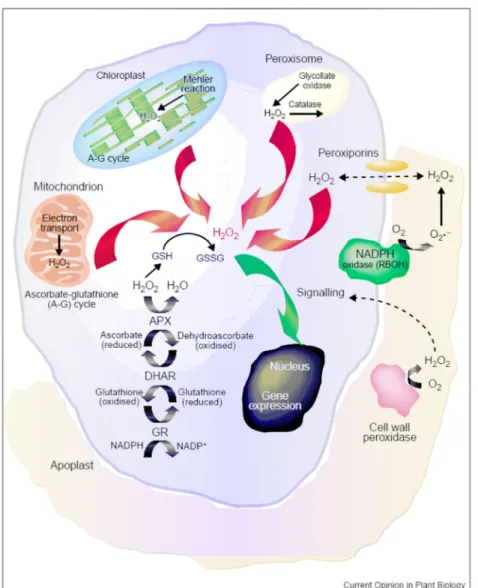
Gli organelli con la maggior attività metabolica ossidativa come cloroplasti, mitocondri e perossisomi, sono i maggiori produttori di ROS nelle cellule vegetali.
Nei cloroplasti, una parte del flusso elettronico, viene indirizzato alla fotoriduzione dell’O2, ridotto ad anione superossido tramite la reazione di Mehler; successivamente, l’O2- viene dismutato ad H2O2 attraverso l’azione delle superossido dismutasi (Cu/Zn-SOD ed Fe-SOD) (Mehler, 1951). Sempre nel cloroplasto, l’H2O2 è il prodotto finale della fotossidazione dell’H2O a livello del fotosistema II (PSII) (Asada, 1999).
Anche i mitocondri sono un sito primario di produzione di ROS, in particolare nei tessuti non fotosintetici e in assenza di luce (Rhoads et al., 2006). In questo caso l’O2- viene generato in due siti specifici della catena di trasporto elettronico, i complessi I e III, e successivamente esso viene convertito ad H2O2 attraverso l’attività della Mn-SOD mitocondriale.
A livello dei perossisomi, la fotorespirazione è strettamente connessa con la produzione di H2O2 che risulta principalmente generata dall’ossidazione del glicolato
ma anche dalla β-ossidazione degli acidi grassi e dall’ossidazione enzimatica di
composti flavinici (Del Rio et al.,1992)
L’H2O2 viene inoltre prodotta, a livelli più bassi, nel citoplasma, nella membrana plasmatica e nella matrice extra-cellulare.
Il sistema ossidasico maggiormente studiato è quello delle NADPH ossidasi di membrana che prove sperimentali, sia di tipo farmacologico che molecolare, hanno individuato come sito di produzione di H2O2, attraverso la formazione di O2-, dovuta al trasferimento di un elettrone dal NADPH all’O2. L’O2- viene poi immediatamente convertito, spontaneamente o attraverso l’azione di SOD, ad H2O2 (Desikan et al., 2003). L’azione della NADPH ossidasi sembra necessitare di ATP extracellulare, come evidenziato nel lavoro di Song et al. (2006) in cui è dimostrato che l’accumulo di O2- in Arabidopsis avviene in seguito a trattamenti esogeni con ATP; al contrario l’overespressione di apirasi (enzima coinvolto nell’idrolisi dei nucleosidi di- e tri- fosfato) ne abbassa i livelli. In effetti, la presenza di un passaggio di ATP dall’interno della cellula verso la matrice sembra avere luogo in presenza di alcuni stress abiotici ma il meccanismo alla base di questo fenomeno non è al momento conosciuto (Jeter et al., 2004). La produzione di H2O2 a livello extracellulare appare come una soluzione ottimale per un meccanismo di signalling tra cellule adiacenti.
Altri risultati, condotti su Arabidopsis, sottolineano l’importante ruolo svolto dalle Rop (Rho-like small G proteins) nella regolazione della produzione di H2O2 attraverso l’enzima NADPH ossidasi. In particolare, bassi livelli nella concentrazione di O2 comporterebbero l’attivazione della via di signalling tramite le proteine Rop che, di conseguenza, attiverebbero l’enzima NADPH ossidasi per la produzione di H2O2.
Fig. 3.1: siti di generazione delle ROS (tratto da Neill et al., 2002b).
3.3 Il ruolo delle ROS in alcuni aspetti della fisiologia della
pianta.
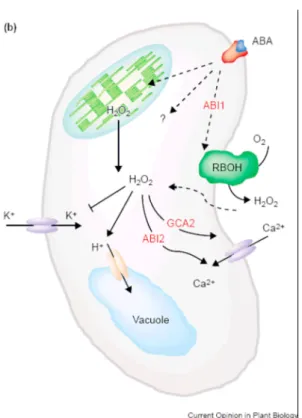
Gli effetti di H2O2 nella chiusura stomatica a livello delle cellule di guardia è noto da tempo in diverse specie vegetali come tabacco e pomodoro (Allan e Flhur, 1997). Nel processo di regolazione della chiusura stomatica, l’acido abscissico (ABA) attiva la sintesi, attraverso l’azione dell’enzima NADPH ossidasi, di H2O2 nelle cellule di guardia. Concentrazioni micromolari di H2O2 sono sufficienti per attivare i canali transmembrana per il flusso di Ca2+ (Pei et al., 2000). Studi successivi (Murata et al., 2001) hanno confermato l’importanza dell’attività dell’enzima NADPH ossidasi in questo processo di regolazione e sono state inoltre individuate due proteine fosfatasi
svolgono un ruolo importante lungo la catena di signalling attivata da ABA e H2O2 (vedi fig.3.2).
Fig. 3.2 (Neill et al., 2002b): ruolo dei geni ABI1 e ABI2 nel signalling dell’ABA sul meccanismo di chiusura stomatica.
Anche NO è stato recentemente individuato come componente della via di
signalling indotta da ABA nelle cellule di guardia, suggerendo, anche in questo caso,
la cooperazione tra H2O2 e NO (Neill et al., 2002a). L’ABA può indurre la sintesi di H2O2 sia da parte dell’enzima di membrana NADPH ossidasi, tramite ABI1, sia da parte del cloroplasto, quest’ultimo osservato soprattutto in Vicia faba (Zhang et al., 2001). Alti livelli intracellulari di H2O2 comportano l’inattivazione dei canali di flusso per K+, provocando l’alcalinizzazione citosolica, e al tempo stesso attivano l’apertura dei canali transmembrana per il flusso di Ca2+, in un’azione mediata da ABI2. Il ruolo delle diverse proteine coinvolte nella via di segnale è stato dedotto grazie a studi effettuati su piante mutanti per questi geni. Quindi è stato osservato che mutanti gca2, (growth control by ABA), rispondono al segnale dell’ABA incrementando la sintesi di H2O2 ma senza attivare l’apertura dei canali di Ca2+. Le cellule di guardia dei mutanti abi1 non portano alla sintesi di H2O2, ma sono in grado
di rispondere ad essa, mentre nel caso dei mutanti abi2 questi possono sintetizzare H2O2 ma non sono in grado di rispondere al suo segnale (Murata et al., 2001).
L’H2O2 risulta essere coinvolta anche nel gravitropismo radicale, mediante l’auxina (Joo et al., 2001). Nelle radici di mais, ad esempio, il gravitropismo è indotto da una produzione asimmetrica di H2O2, esattamente come avviene nel caso di produzione asimmetrica delle auxine. Altri studi hanno dimostrato che l’applicazione asimmetrica di H2O2 o di antiossidanti promuove o inibisce, rispettivamente, la risposta gravitropica. Infatti, la presenza di catalasi non ha alcun effetto sulla risposta gravitropica, suggerendo che l’H2O2 sia coinvolta. Il meccanismo di generazione di H2O2 in questo tipo di risposta resta al momento ancora sconosciuto (Neill et al., 2002b).
Da studi effettuati su mutanti recessivi di Arabidopsis che mostrano un alterato fenotipo dei peli radicali, è stato osservato come sia la produzione che la localizzazione delle ROS fanno parte dei meccanismi di crescita che determinano la forma di queste cellule specializzate (Carol e Dolan, 2006). I peli radicali del mutante di Arabidopsis rhd2 (root hair defective 2) non sono infatti in grado di allungarsi. Il gene mutato RHD2 codifica per una respiratory burst oxidase
homologue C (AtrbohC), ovvero una NADPH ossidasi di membrana. Il mutante rhd2
produce quindi livelli dimezzati di ROS rispetto al suo background, il quale mostra inizialmente un accumulo fisiologico di ROS a livello del rigonfiamento del tricoblasto e, durante l’allungamento, nella porzione terminale del pelo. Nel caso del mutante rhd2 questo accumulo non è visibile o è estremamente ridotto e questo fenotipo sembra quindi essere ricollegabile alla mancata produzione di ROS da parte di RHD2 (Foreman et al., 2003).
3.4 Meccanismi di trasduzione del segnale mediati da ROS
La produzione di H2O2 risulta incrementata in risposta a vari tipi di stress, quali carenza idrica, alte e basse temperature, stress termico e luminoso (Neill et al., 2002a), per questo motivo risulta assai probabile che essa possa essere un fattore chiave in fenomeni di acclimatazione e di tolleranza.
Generalmente la produzione di H2O2 viene bilanciata da efficienti sistemi antiossidanti (Noctor e Foyer, 1998; Corpas et al., 2001), ma, in presenza di stress abiotici e biotici, questa omeostasi viene alterata, culminando nell’attivazione di
meccanismi di signalling per il controllo di processi di risposta specifici quali difesa, crescita, sviluppo attraverso l’alterazione dell’espressione genica.
In seguito all’accumulo di ROS, l’alterazione dello stato redox di ogni compartimento cellulare può determinare una differente regolazione di alcune proteine, in particolare quelle coinvolte nei meccanismi di fosforilazione a cascata e nei flussi del Ca2+ (Nakagami et al., 2006; Neill et al., 2002a).
Da un punto di vista chimico, H2O2 è una molecola estremamente semplice, per cui difficilmente esistono specifici recettori in grado di legarsi ad essa per attivare una via di signalling, mentre è assai probabile che, a causa della sua natura debolmente ossidante, sia in grado di interagire con i residui di cisteina, ma anche di tirosina, triptofano e istidina delle proteine (Vranova et al., 2002). Affinché però queste interazioni abbiano un significato fisiologico, dovranno indurre dei cambiamenti conformazionali che alterino l’attività della proteina e che siano sufficientemente duraturi da poter attivare la risposta cellulare (Neill et al., 2002).
Analisi microarray sono state utilizzate per verificare la suscettibilità del trascrittoma ad applicazioni esogene di H2O2 sia in Arabidopsis che in altre specie vegetali (Gadjev et al., 2006; Kim et al., 2005; Vandenbroucke et al., 2008; Desikan
et al., 2001a ) e sono stati individuati diversi geni che possono essere indotti o
repressi. L’espressione di tutti questi geni varia in risposta a differenti stress abiotici e biotici sia in maniera direttamente dipendente che indipendente da H2O2, suggerendo la presenza di una vera e propria interazione nelle diverse vie di
signalling (Desikan et al., 2001). Da queste analisi trascrittomiche è stata individuata
inoltre una generale induzione da parte di H2O2 di geni coinvolti nel signalling dell’etilene, tra cui alcune istidine chinasi. Uno studio più approfondito è stato quindi condotto sul recettore istidina chinasi ETR1 di Arabidopsis, essenziale per la percezione di H2O2 nel processo che porta alla chiusura degli stomi (Desikan et al., 2005). Questa affinità di legame è dovuta alla cisteina localizzata in posizione 65 (Cys65), in quanto nel mutante etr1-1, in cui la Cys65 è sostituita dalla tiroxina, la risposta all’ H2O2 nella chiusura stomatica è assente. Nel mutante etr1-3, invece, alterato nell’attività del dominio dell’istidina chinasi, l’attività di signalling di H2O2 è mantenuta, mentre la percezione dell’etilene risulta persa. Questo suggerisce non solo che l’attività di signalling da parte di H2O2 non sia correlata all’attività dell’isitidina chinasi, ma anche che il signalling via ETR1 indotto da H2O2 sia separato da quello indotto dall’etilene (Hancock et al., 2006).
Studi sul mutante TM219 di S. cerevisiae, estremamente suscettibile alla presenza di H2O2 per la mancanza del sistema funzionale a due componenti SLN1-SSK1, hanno mostrato che la trasformazione con il gene ETR1 di Arabidopsis ristabilisce la tolleranza a H2O2 (Singh, 2000) della quale è responsabile, anche in questo caso, la Cys65(Desikan etal., 2005).
La molecola H2O2 è in grado, inoltre, di attivare il segnale a cascata delle MAP-kinasi nei vari tessuti (Desikan et al., 2001), ma come questa attivazione avvenga non è ancora noto. L’attivazione costitutiva del meccanismo H2O2-MAPK è in grado di dare tolleranza a diversi stress ambientali (Kovtun et al., 2000).
Le “mitogen-activated protein kinase kinase kinase” (MAPKKKs) svolgono un ruolo chiave nelle vie di signalling intra- ed extra-cellulari. Rappresentano, infatti, importanti regolatori di crescita, differenziamento, proliferazione e risposta a diversi tipi di stress (Garrington e Johnson, 1999). Il meccanismo cosiddetto “a cascata” di queste proteine consiste in una modulazione di tipo MAPKKK-MAPKK-MAPK che suggerisce l’associazione a specifici recettori di H2O2 che si trovano a monte della cascata di signalling e a molecole target a valle. I recettori possono attivare le MAPKKK o attraverso un’interazione di tipo fisico o tramite fosforilazione da parte dello stesso recettore (Nakagami et al., 2006). Tra i possibili target ci possono essere sia fattori di trascrizione ma anche altre proteine kinasi. Studi recenti (Nakagami et
al., 2004) hanno portato all’isolamento di una oxidative stress-activated MAP triple
kinase 1 (OMTK1) in Medicago sativa. Tra le varie sostanze testate questa MAPKKK risulta essere attivata solo da H2O2. Una simile MAPKKK è stata individuata anche in Arabidopsis (Nakagami et al., 2006). Si tratta di MEKK1 implicata nei meccanismi di segnale a cascata durante stress abiotici e biotici. In particolare, MEKK1 è coinvolta nell’attivazione di alcune MAPKinasi che svolgono un ruolo cruciale nell’adattamento a stress abiotici e nei meccanismi di difesa da agenti patogeni, come MPK3, MPK4 e MPK6. Studiando un mutante inserzionale
mekk1 è stato dimostrato che MEKK1 è un attivatore di MPK4, mentre rappresenta