The Association of Palmitoylethanolamide with Luteolin
Decreases Autophagy in Spinal Cord Injury
Rosalba Siracusa1&Irene Paterniti1&Giuseppe Bruschetta1&Marika Cordaro1& Daniela Impellizzeri1&Rosalia Crupi1&Salvatore Cuzzocrea1,2&Emanuela Esposito1
Received: 9 March 2015 / Accepted: 26 June 2015 / Published online: 5 July 2015 # The Author(s) 2015. This article is published with open access at Springerlink.com
Abstract Spinal cord injury (SCI) is a devastating condition of the central nervous system (CNS) often resulting in severe functional impairment and for which there are not yet restor-ative therapies. In the present study, we performed a widely used model of SCI to determine the neuroprotective propriety of palmitoylethanolamide (PEA) and the antioxidant effect of a flavonoid luteolin (Lut), given as a co-ultramicronized com-pound co-ultraPEALut. In particular, by western blot analysis and immunofluorescence staining, we investigated whether this compound (at the dose of 1 mg/kg) was able to modulate autophagy. Our results showed that treatment with co-ultraPEALut after SCI reduced the expression of proteins pro-moter of autophagy such as Beclin-1 and microtubule-associated protein 1A/1B-light chain 3 (MAP-LC3). In con-trast, this compound decreased the levels of mammalian target of rapamycin (mTOR), p-Akt, and p-70S6K which are pro-teins that inhibit autophagy. These data confirmed that the protective role of co-ultraPEALut is associated with inhibition of excessive autophagy and regulation of protein degradation. Therefore, treatment with co-ultraPEALut could be consid-ered as a possible therapeutic approach in an acute traumatic lesion like SCI.
Keywords Neuroprotection . Anti-inflammatory . Anti-oxidant . Autophagy . Spinal compression
Introduction
Spinal cord injury (SCI) is defined as an acute traumatic lesion of neural elements in the spinal canal (spinal cord and cauda equina), resulting in a change, either temporary or permanent, in normal motor, sensory, or autonomic function. Spinal cord injury usually begins with a sudden, traumatic blow to the spine that causes local segmental damage to the spinal cord, which is called primary injury [1,2]. The primary damage to the tissue is followed by a second phase of tissue degenera-tion, the secondary injury that can occur over weeks or even months. In secondary injury, acute inflammation can develop into a chronic process if feedback mechanisms fail to inhibit amplification of the inflammatory response. Chronic inflam-mation leads to a continuous influx of neutrophils, macro-phages, lymphocytes, and eosinophils from the circulation, causing more destruction and scarring of tissue [3]. Cell death resulting from all of these mechanisms occurs through necrot-ic and apoptotnecrot-ic phenomena or autophagy.
Concern about autophagy has recently increased because of its potential role in neurodegenerative diseases, such as Parkinson’s and Alzheimer’s diseases, where autophagy may be a protective mechanism [4]. Earlier studies also demon-strated increased autophagic activity at lesion sites after cere-bral hypoxia-ischemia (HI) injury [5], intracerebral hemor-rhage (ICH) [6], and traumatic brain injury (TBI) [7,8].
Autophagy, literallyBself-eating, is now recognized as an intracellular catabolic mechanism for the degradation, elimi-nation, and recycling of long-lived proteins and unwanted organelles in a cell during development and under stress con-ditions [9,10]. Therefore, autophagy plays an important role
* Emanuela Esposito [email protected]
1
Department of Biological and Environmental Sciences, University of Messina, Viale Ferdinando Stagno D’Alcontres, 31,
98166 Messina, Italy
2 Department of Pharmacological and Physiological Science, Saint
Louis University School of Medicine, 1402 South Grand Blvd, St Louis, MO 63104, USA
in homeostasis and also to a kind of cell death, which is known as autophagic cell death [11,12]. It begins with the formation of double-membraned vesicle that subsequently engulfs cyto-plasmic components, including cytosolic proteins and organ-elles, to become autophagosomes (APs). APs fuse with lyso-somes to form autolysolyso-somes, and intra-autophagosomal components are degraded by lysosomal hydrolases. Several key molecular components participate in the initiation, pro-gression, and completion of autophagy, like the mammalian target of rapamycin (mTOR) that inhibits autophagy, whereas Beclin 1 and LC3 promote it [8,13]. However, autophagy seems to be a double-edged sword that protects cells when it is moderately activated, yet its excessive activation can induce autophagic cell death [14].
In recent years, the pathophysiology of SCI has been a focus of extensive studies; animal models have been proved to be important tools in this field and are employed to inves-tigate the mechanisms of primary and secondary injury. Pre-vious studies have shown that autophagy plays a key role in secondary injury in both animal models and human tissue by causing progressive degeneration of the spinal cord [15,16]. However, the molecular pathway of secondary injury and the role of autophagy in the recovery of SCI remain unclear. Fol-lowing acute spinal cord injury, an increase in mTOR expres-sion and p70S6K activity also may be required for functional improvement [17]. The involvement of autophagic mecha-nism underlying spinal neuroprotective effects of palmitoylethanolamide (PEA) and luteolin association was never been investigated. Thus, we studied the autophagic pathway involved in spinal cord compression and to explore the effects of the treatment with a new co-ultramicronized composite of N-PEA and luteolin (Lut) on autophagy expres-sion. This formulation (co-ultraPEALut) is based on associa-tion of anti-inflammatory PEA with anti-oxidant Lut on a 10:1 mass basis. We have previously demonstrated that treatment with co-ultraPEALut significantly reduced the inflammatory process in an ex vivo model of spinal cord organotypic cul-tures and in experimental mouse models of TBI [18,7]. Fur-thermore in this last model, we showed that co-ultraPEALut reduced autophagy after TBI [7].
Therefore, the aim of this study was to demonstrate that treatment co-ultraPEALut inhibited autophagy even after SCI, having neuroprotective effect.
Methods
Animals
CD1 mice (male 25–30 g; Harlan Nossan, Milan, Italy) were housed in a controlled environment and provided with stan-dard rodent chow and water. Mice were housed in stainless steel cages in a room kept at 22±1 °C with a 12-h light, 12-h
dark cycle. The animals were acclimatized to their environ-ment for 1 week and had ad libitum access to tap water and rodent standard diet. The study was approved by the Univer-sity of Messina Review Board for the care of animals. All animal experiments complied with regulations in Italy (D.M. 116192) as well as the EU regulations (O.J. of E.C. L 358/1 12/18/1986).
Co-ultramicronization Process of PEA and Lut
The co-ultramicronization process was performed using jet mill equipment (Sturtevant, Inc., 348 Circuit Street Hanover, MA, USA) with a chamber of 300 mm in diameter, operated withBspiral technology^ and driven by compressed air at 10 to 12 bars. Crashing was determined by the high number of collisions that occurred among particles as a result of the high level of kinetic (not mechanical) energy. This process is effec-tive not only in reducing product particle size but also in modifying crystalline structure. Scanning electron microscopy showed an intimate intermixing of PEA and Lut, while anal-ysis by differential scanning calorimetry and X-ray diffraction indicated transformation into a crystalline form different from the original two, definable asBa higher energy content form.^ The composite showed the following particle size distribution: 96 % <10 μm, 80 % <5 μm, and 40 % <2 μm. Co-ultraPEALut (Epitech Group s.r.l.) was dissolved in 10 % eth-anol (Sigma-Aldrich, St Louis, MO, USA) and used at a con-centration of 1 mg/kg in the in vivo study.
The dose of co-ultraPEALut (1 mg/kg) used was based on previous in vivo studies [18].
SCI
Mice were anesthetized with intraperitoneal administration of ketamine and xylazine (2.6 and 0.16 mg/kg body weight, re-spectively). A longitudinal incision was made on the midline of the back, exposing the paravertebral muscles, as previously described [18]. These muscles were dissected away, the spinal cord was exposed via a four-level T5 to T8 laminectomy, and SCI was produced by extradural compression at T6 to T7 level, using an aneurysm clip with a closing force of 24 g. Following surgery, 1.0 cm3of saline was administered subcu-taneously in order to replace the blood volume lost during the surgery. During recovery from anesthesia, mice were placed on a warm heating pad and covered with a warm towel. The mice were individually housed in a temperature-controlled room at 27 °C. Food and water were provided to the mice ad libitum. During this time period, the animals’ bladders were manually voided twice a day until the mice were able to regain normal bladder function. In all injured groups, the spinal cord was compressed for 1 min. Sham animals were only subjected to laminectomy. Spinal cord tissues were taken at 24 h after trauma. Tissue segments contained the lesion
(1 cm on each side of the lesion), according to counts of retrogradely labeled neurons at the origin of distinct descend-ing motor pathways and to morphometric assessments of nor-mal residual tissue at the injury epicenter, as previously de-scribed [19].
Experimental Groups and Treatments
Mice Were Randomly Allocated into the Following Groups
1. Sham+vehicle: mice were subjected to laminectomy, but the aneurysm clip was not applied and treated intraperito-neally (i.p.) with vehicle (n=30).
2. Sham+co-ultraPEALut (10:1)=same as the Sham+vehi-cle group, but co-ultraPEALut was administered at the final dose of 1 mg/kg i.p. at 1 and 6 h after laminectomy (n=30).
3. SCI+vehicle: mice were subjected to laminectomy and the aneurysm clip was applied (n=30).
4. SCI+co-ultraPEALut (10:1): mice were subjected to SCI and administered co-ultraPEALut at the final dose of 1 mg/kg i.p. at 1 and 6 h after SCI (n=30).
Western Blot Analysis
Cytosolic and nuclear extracts were prepared with slight mod-ifications of a published procedure [20]. Spinal cord tissue from each mouse was suspended in extraction Buffer A con-taining 0.2 mM PMSF, 0.15 mM pepstatin A, 20 mM leupeptin, 1 mM sodium orthovanadate, homogenized at the highest setting for 2 min, and centrifuged at 12,000 rpm for 4 min at 4 °C. Supernatants represented the cytosolic fraction. The pellets, containing enriched nuclei, were resuspended in Buffer B containing 1 % Triton X-100, 150 mM NaCl, 10 mM Tris–HCl pH 7.4, 1 mM EGTA, 1 mM EDTA, 0.2 mM PMSF, 20 mm leupeptin, and 0.2 mM sodium orthovanadate. After centrifugation 10 min at 12,000 rpm at 4 °C, the supernatants that we obtained were the nuclear proteins. Protein concentrations were estimated by the Bio-Rad protein assay using bovine serum albumin as standard. Briefly, samples were heated to 100 °C for 5 min, and equal amounts of protein were separated on 18 % SDS-PAGE gel and transferred to nitrocellulose mem-brane. Specific primary antibody, anti- anti-MAPLC3α (1:500, Santa Cruz Biotechnology), Beclin-1 (1:500; Santa Cruz Biotechnology), anti-p62 (1:1000; Cell Signaling), anti-mTOR (1:1000; Cell Signaling), anti-p70S6K (1:1000; Cell Signaling), and p-Akt (1:1000; Cell Signaling) were mixed in 1× PBS, 5 % w/v nonfat dried milk, 0.1 % Tween-20 (PMT), and incubated at 4 °C, overnight. After, mem-branes were incubated with peroxidase-conjugated bovine anti-mouse IgG secondary antibody or peroxidase-conjugated goat anti-rabbit IgG (1:2000, Jackson
ImmunoResearch) for 1 h at room temperature. To ascertain that blots were loaded with equal amounts of protein lysates, they were also incubated in the presence of the antibody againstβ-actin (1:5000; Santa Cruz Biotechnology). Signals were detected with enhanced chemiluminescence detection system reagent according to manufacturer’s instructions (SuperSignal West Pico Chemiluminescent Substrate, Pierce). The relative expression of the protein bands was quantified by densitometry with Bio-Rad ChemiDoc™ XRS+software and standardized toβ-actin levels. Images of blot signals (8-bit/ 600-dpi resolution) were imported to analysis software (Image Quant TL, v2003). A preparation of commercially available molecular weight markers made of proteins of molecular weight 10 to 250 kDa was used to define molecular weight positions and as reference concentrations for each molecular weight.
Immunofluorescence Staining
After deparaffinization and rehydration, detection of GFAP, NeuN, Beclin-1, and MAPLC3-α was carried out after boiling in 0.1 M citrate buffer for 1 min. Nonspecific adsorption was minimized by incubating the section in 2 % (v/v) normal goat serum in PBS for 20 min. Sections were incubated with mouse monoclonal anti-NeuN (1:100, v/v Millipore), or with mouse monoclonal anti-GFAP (1:100, v/v Santa Cruz Biotechnolo-gy), or with polyclonal rabbit anti-Beclin-1 (1:100, v/v Santa Cruz, Biotechnology), or with rabbit anti-MAPLC3α (1:100, v/v Santa Cruz Biotechnology) antibody in a humidified chamber for O/N at 37 °C. Sections were washed with PBS and were incubated with secondary antibody FITC-conjugated anti-mouse Alexa Fluor-488 antibody (1:2000 v/v Molecular Probes, UK) and with TEXAS RED-conjugated anti-rabbit Alexa Fluor-594 antibody (1:1000 in PBS, v/v Molecular Probes, UK) for 1 h at 37 °C. Sections were washed and for nuclear staining 4′,6′-diamidino-2-phenylindole (DAPI; Hoechst, Frankfurt; Germany) 2μg/ml in PBS was added. Sections were observed and photographed at ×20 magnification using a Leica DM2000 microscope (Leica). All images were digitalized at a resolution of 8 bits into an array of 2560×1920 pixels. Optical sections of fluores-cence specimens were obtained using a HeNe laser (543 nm), a laser UV (361–365 nm), and an argon laser (458 nm) at a 1-min, 2-s scanning speed with up to 8 averages; 1.5μm sections were obtained using a pinhole of 250. Contrast and brightness were established by examining the most brightly labeled pixels and applying settings that allowed clear visualization of struc-tural details while keeping the highest pixel intensities close to 200. The same settings were used for all images obtained from the other samples that had been processed in parallel. Digital images were cropped and figure montages prepared using Adobe Photoshop CS5 (Adobe Systems; Palo Alto, CA).
Light Microscopy
For histopathological examination by standard hematoxylin and eosin (H&E) staining, the mice were anesthetized with ketamine and xylazine (2.6 and 0.16 mg/kg body weight respectively) 24 h after injury and then perfused transcardially with cold PBS (0.1 M). Tissues were removed under magnified vision, and segments containing the lesion (1 cm on each side of the lesion) were collected in 4 % paraformaldehyde for proper fix-ation and then dehydrated with graded ethanol and embedded in paraffin wax. Sections of 5-μm thickness were cut into longitu-dinal sections for the posterior area of the spinal cord, stained with H&E, and studied using light microscopy (Dialux 22 Leitz). Representative images were shown. Blinded histologic scoring (Fig.1d) was performed on a 6-point scale: 0, normal; 1, 1–5 eosinophilic neurons within the gray matter area; 2, 5–10 eosinophilic neurons; 3, >10 eosinophilic neurons; 4, a small infarction (less than one-third of the gray matter area); 5, a mod-erate infarction (one-third to one-half of the gray matter area); and 6, a large infarction (more than half of the gray matter area).
Materials
Unless otherwise stated, all compounds were obtained from Sigma-Aldrich, while co-ultraPEALut was a kind gift from Epitech Group s.r.l. (Saccolongo, Italy). All other chemicals were of the highest commercial grade available. All stock solutions were prepared in non-pyrogenic saline (0.9 % NaCl, Baxter, Milan, Italy) or 10 % dimethyl sulfoxide.
Statistical Evaluation
All values in the figures and the text are expressed as mean± SEM. Results shown in the figures are representative of at least three experiments performed on different in vivo exper-imental days. In each experiment, we used five animals per group, unless otherwise indicated. The results were analyzed by one-way analysis of variance followed by a Bonferroni post hoc test for multiple comparisons. A p value of less than 0.05 was considered significant.
Fig. 1 Effects of co-ultraPEALut treatment on histological alterations of the spinal cord tissue 24 h after injury. Compared with the Sham-surgery mice (a, a1), significant histologic damage to the spinal cord was evident in mice subjected to SCI and treated with vehicle (b, b1). In the perilesional area of the SCI mice, edema was evident, as and alteration
of the white matter. A significant reduction of SCI-associated damage was observed in the tissue samples collected from mice treated with co-ultraPEALut (c, c1). These data are also visible in the histological score (d). ***p<0.001 vs Sham; ###p<0.001 vs SCI
Results
Effect of Co-ultraPEALut on Histological Parameters
To evaluate the severity of the trauma at the level of the perilesional area at 24 h after injury, the sections obtained from each group were stained with hematoxylin and eosin (H&E). Compared with the sham group (Fig.1a, b, see mag-nification higher a1 and b1, respectively), the SCI+vehicle group showed the presence of edema and alteration of the white matter (Fig.1b, d, see magnification higher b1). Treat-ment with co-ultraPEALut attenuated acute inflammation and injury at 24 h after SCI (Fig.1c, d, see magnification higher c1).
Effect of Co-ultraPEALut on Autophagy After SCI: Expression of Beclin-1, p62, and MAP-LC3
To investigate the mechanisms of autophagy after SCI, we examined levels of proteins involved in the regulation and formation of autophagosomes. Beclin-1 has a central role in autophagy because it interacts with various cofactors inducing
autophagy. To compare the level of Beclin-1 protein between the injured and the treated spinal cord section, western blot analysis was performed. We showed that Beclin-1 expression dramatically increased in the spinal cord tissue (T6 to T8 tract) collected from mice subjected to SCI compared to the sham group, while the treatment with co-ultraPEALut significantly reduced SCI-induced Beclin-1 expression (Fig.2a, see densi-tometry analysis in a1). For more accurate quantification of co-ultraPEALut effects on autophagy process, we examined levels of p62 by western blot analysis. We detected that p62 expression increased after SCI, whereas the treatment with co-ultraPEALut significantly reduced levels of this protein (Fig.2b, see densitometry analysis in b1). p62 also interacts with a central component of the machine autophagy, autoph-agic marker MAP-LC3, and carries the altered proteins to degradation by autophagy. Therefore, by western blot analy-sis, we showed that the ratio of LC3 I to LC3 II increased in untreated animals 24 h after injury, though this ratio signifi-cantly diminished to near sham levels of expression following co-ultraPEALut treatment (Fig.2c, see densitometry analysis in c1), suggesting a reversal of autophagic activity upregula-tion seen following SCI.
Fig. 2 Effect of co-ultraPEALut about proteins promoting autophagy after SCI. Western blot analysis for Beclin-1, p62, and LC3 showed an increase of levels of these proteins in SCI group compared to the Sham group, whereas the treatment with co-ultraPEALut significantly reduced
SCI-induced Beclin-1, p62, and LC3 expression (a, a1; b, b1; c, c1 respectively). The data are representative of at least three independent experiments. a **p<0.01 vs Sham; ##p<0.01 vs SCI. b **p<0.01 vs Sham; #p<0.05 vs SCI. c *p<0.05 vs Sham; #p<0.05 vs SCI
Effect of Co-ultraPEALut on Beclin-1 and MAP-LC3 Expression by Immunofluorescence Staining
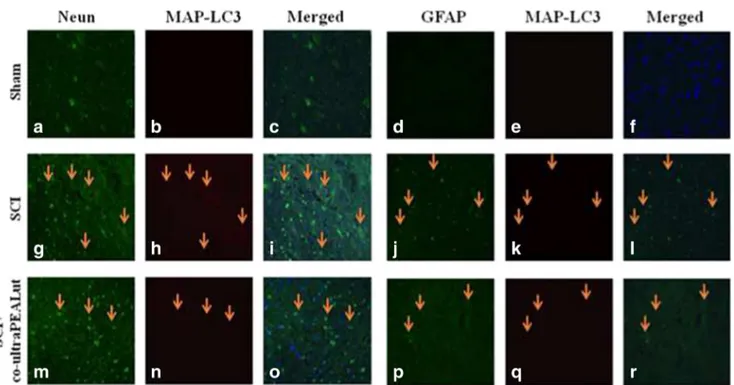
To better understand the Beclin-1 and MAP-LC3 expression in a specific population of cells, including neurons and astrocytes, the spinal cord sections (T6 to T8 tract) at 24 h after compres-sion were double-stained for Beclin-1 (red) or MAP-LC3 (red) and various cell type markers, such as NeuN (green) for neurons and GFAP (green) for astrocytes (Figs.3and4). In the double staining, the expression of Beclin-1 was observed in NeuN and GFAP-labeled cells of the injured SCI group (Fig.3g–iandj–l, respectively) compared to Sham (Fig.3a–candd–f, respective-ly), whereas a treatment with co-ultraPEALut not only de-creased significantly these proteins expression but also the dou-ble staining is less observed in both NeuN and GFAP (Fig.3m– oandp–r, respectively). Similarly, at 24 h after SCI, double staining of LC3 revealed a strong co-localization with NeuN and GFAP in SCI group (Fig.4g–iandj–l, respectively) com-pared to the Sham group (Fig.4a–candd–f, respectively), while treatment of co-ultraPEALut decreased significantly these pro-tein expressions (Fig.4m-oandp-rrespectively).
Effect of Co-ultraPEALut on PI3K/Akt/mTOR Pathway After SCI
The PI3K/Akt/mTOR pathway is an intracellular signaling pathway important in regulating the cell cycle. mTOR is
upstream signal in the regulation of autophagy which is pos-itively regulated by PI3K/Akt and results in the inhibition of autophagy. Therefore, by western blot analysis, we detected the activity of Akt/mTOR signaling pathways in our model in vivo. Our results showed that p-Akt and mTOR levels de-creased significantly after SCI, compared with the Sham group, while co-ultraPEALut treatment increased p-Akt and mTOR expression, inhibiting autophagy (Fig.5a, c, see den-sitometry analysis in a1 and b1, respectively).
Moreover, a downstream target of mTOR signal a p70S6K activity was assessed by western blot analysis. p70S6k activ-ity was suppressed in mice after SCI, whereas the co-ultraPEALut treatment enhanced activation of p70S6K signal-ing (Fig.5c, see densitometry analysis in c1).
Discussion
The CNS is sensitive to mechanical injuries causing perma-nent functional deficits such as the case of patients who have SCI. The mechanical forces imparted to the spinal cord cause immediate tissue disruption, with direct axonal and neuronal injury, inducing the death of neurons. Moreover, neurons con-tinue to die for hours or days after SCI, as a consequence of excitotoxicity, vascular dysfunction, and, in particular, be-cause of the ensuing inflammatory response [21]. Inflamma-tory responses are a major component of secondary injury and
Fig. 3 Co-localization of Neun/Beclin-1 and GFAP/Beclin-1 after SCI. Results are shown for a–f Sham group, g–l mice with SCI, and m–r mice with SCI treated with co-ultraPEALut. Double-stained sections at 24 h post injury indicated that Beclin-1 expression increased in both neurons and astrocytes respectively. Orange arrows indicate co-localizations (g–l)
and revealed a high co-localization between NeuN/Beclin1 and GFAP/ Beclin-1 double staining. Treatment with the co-ultraPEALut reduced Beclin-1 expression (m–r). All images were digitalized at a resolution of 8 bits into an array of 2048×2048 pixels
play a central role in regulating the pathogenesis of acute and chronic SCI [22]. Functional recovery after SCI is often poor and, as yet, there are no nonsurgical therapies that have been successfully translated for use in the clinic. Therefore, there is a great need for drugs that will reduce post-traumatic, inflammation-related spinal cord damage because it has been reported that reducing inflammation decreases secondary de-generation and the functional deficit after SCI. In a previous study, we showed that co-ultraPEALut treatment (1 mg/kg) exerted beneficial effects in ex vivo model using spinal cord organotypic slice cultures and in in vivo model of spinal cord compression injury in mice. Briefly, we found that co-ultraPEALut had anti-inflammatory, antioxidant, and neuro-protective properties [18].
Furthermore, recent studies demonstrated that another pos-sible consequence of SCI is energy depletion, and this elicits autophagy [15,16,23]. Autophagy is a cellular catabolic deg-radation response to starvation or stress, whereby cellular pro-teins, organelles, and cytoplasm are engulfed, digested, and recycled to sustain cellular metabolism [9,24]. Constitutive, basal autophagy also has an important homeostatic function, by maintaining protein and organelle quality control. Al-though most evidence supports a role for autophagy in sus-taining cell survival, paradoxically, cell death resulting from progressive cellular consumption has been attributed to unre-strained autophagy [25–27]. The mechanisms that regulate the mutually opposed survival and death roles for autophagy are
still unknown. The most plausible explanation is that catabo-lism through autophagy is predominantly survival-supporting, but that an imbalance in cell metabolism, where autophagic cellular consumption exceeds the cellular capacity for synthe-sis, promotes cell death [28].
Therefore, the aim of the present study was to explore new therapeutic approaches promoting survival of cells. In partic-ular, we wanted to investigate whether the new compound co-ultraPEALut (at the dose of 1 mg/kg) was able to modulate autophagy promoting functional recovery after SCI.
In the first step, using H&E staining, we evaluated the effect of co-ultraPEALut on spinal cord at the level of the perilesional area at 24 h after injury. Histological analysis showed that mice, after SCI, presented edema and alteration of the white matter, while mice treated with co-ultraPEALut showed an attenuation of acute inflammation and injury after SCI.
In the second step, we studied the effects of this new com-pound on autophagy markers after SCI. At first, we examined the levels of proteins that promote autophagy such as Beclin-1 and MAP-LC3. Beclin-1 is a protein that participates in the regulation of autophagy and has an important role in develop-ment, tumorigenesis, and neurodegeneration [29]. Beclin-1 functions as a scaffold for the formation of the PI3K complex, one of the first components recruited during the development of autophagosomes. During autophagy initiation and autophagosome formation, Beclin-1 binds
microtubule-Fig. 4 Co-localization of NeuN/LC3 and GFAP/LC3 after SCI. Results are shown for (a–f) Sham group, (g–l) mice with SCI, and (m–r) mice with SCI treated with co-ultraPEALut. Double-stained sections at 24 h post-injury indicated that LC3 expression increased in both neurons and astrocytes respectively. Orange arrows indicate co-localizations (g–l) and
revealed a high co-localization between NeuN/LC3 and GFAP/LC3 double staining. Treatment with the Co-ultraPEALut reduced Beclin-1 expression (m–r). All images were digitalized at a resolution of 8 bits into an array of 2048×2048 pixels
associated protein 1A/1B-light chain 3 I (LC3I) that is con-verted to its membrane-bound form (LC3II), which then inter-acts with the ubiquitin-binding protein p62/sequestosome 1 [30–32]. Since p62 accumulates when autophagy is inhibited and decreased levels can be observed when autophagy is in-duced, p62 may be used as a marker to study autophagic flux [32]. In this study, we observed an increase of the expression of Beclin-1, LC3, and p62 after SCI that dramatically de-creased after treatment with co-ultraPEALut. Furthermore, we wanted to assess whether Beclin-1 and MAP-LC3 were expressed in specific cells. Therefore, the spinal cord sections at 24 h after compression were double-stained for Beclin-1 or MAP-LC3 and various cell type markers, such as NeuN for neurons and GFAP for astrocytes. By immunofluorescence staining, we observed a strong co-localization of Beclin-1 and LC3 with Neun and GFAP. These results showed that after SCI, the expression of these two proteins increases in neurons and astrocytes, while treatment with co-ultraPEALut reduces the levels of Beclin-1 and LC3 in both cell types.
Finally, we wanted to evaluate the effect of co-ultraPEALut on Akt/mTOR/p70S6K pathway that in autophagy has been considered a central regulatory pathway of the protein trans-lation involved in regulating cell proliferation, growth, differ-entiation, and survival [33,34]. mTOR kinase is a master regulator of autophagy directly suppressing autophagy. This protein is a central signal integrator that functions as a check-point with upstream Akt and downstream p70S6K being the two most important mediators [8]. Therefore, using western blot analysis, we detected mTOR expression and the phos-phorylation of Akt after SCI. Our results indicated that p-Akt and mTOR levels decreased significantly after SCI, while co-ultraPEALut treatment increased expression of these pro-teins. Moreover, p70S6k activity was suppressed in mice after SCI, whereas the co-ultraPEALut treatment enhanced activa-tion of p70S6K signaling. These data confirmed that the pro-tective role of co-ultraPEALut is associated with inhibition of excessive autophagy via Akt/mTOR/p70S6K signals and reg-ulation of protein degradation.
Fig. 5 Effect of co-ultra PEALut about proteins inhibiting autophagy after SCI. Western blot analysis for mTOR, p-Akt, and p70S6K showed a decrease of levels of these proteins in SCI group compared to the Sham group, whereas the treatment with co-ultraPEALut significantly increased
SCI-induced mTOR, p-Akt, and p70S6K expression (a, a1; b, b1; c, c1 respectively). The data are representative of at least three independent experiments. a **p<0.01 vs Sham; #p<0.05 vs SCI. b **p<0.01 vs Sham; ##p<0.01 vs SCI. c ***p<0.001 vs Sham; ##p<0.01 vs SCI
We conclude that co-ultraPEALut treatment at the dose of 1 mg/kg is capable to improve neurobehavioral functions, and reduce apoptotic cell death and neuroinflammation, improv-ing tissue structure after SCI [18]. In addition, our results suggest that SCI induces autophagic cell death at the lesion site; therefore, co-ultraPEALut treatment inhibiting autophagy in in vivo model of SCI has protective effect. In conclusion, co-ultraPEALut could be considered as a possible therapeutic target in an acute traumatic lesion like SCI.
Acknowledgments The authors would like to thank Maria Antonietta Medici for her excellent technical assistance during this study and Mr Francesco Soraci for his secretarial and administrative assistance and Miss Valentina Malvagni for her editorial assistance with the manuscript. This study was supported by PON 01_02512.
Compliance with Ethical Standards
Conflict of Interest The authors declare that they have no conflict of interest.
The study was approved by the University of Messina Review Board for the care of animals. All animal experiments complied with regulations in Italy (D.M. 116192) as well as the EU regulations (O.J. of E.C. L 358/1 12/18/1986).
Open Access This article is distributed under the terms of the Creative C o m m o n s A t t r i b u t i o n 4 . 0 I n t e r n a t i o n a l L i c e n s e ( h t t p : / / creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appro-priate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
References
1. Davalos D, Grutzendler J, Yang G, Kim JV, Zuo Y, Jung S, Littman DR, Dustin ML et al (2005) ATP mediates rapid microglial re-sponse to local brain injury in vivo. Nat Neurosci 8(6):752–758 2. Amar AP, Levy ML (1999) Pathogenesis and pharmacological
strategies for mitigating secondary damage in acute spinal cord injury. Neurosurgery 44(5):1027–1039, discussion 1039-1040 3. Bareyre FM, Schwab ME (2003) Inflammation, degeneration and
regeneration in the injured spinal cord: insights from DNA micro-arrays. Trends Neurosci 26(10):555–563
4. Nixon RA, Cataldo AM, Mathews PM (2000) The endosomal-lysosomal system of neurons in Alzheimer's disease pathogenesis: a review. Neurochem Res 25(9-10):1161–1172
5. Carloni S, Buonocore G, Balduini W (2008) Protective role of autophagy in neonatal hypoxia-ischemia induced brain injury. Neurobiol Dis 32(3):329–339. doi:10.1016/j.nbd.2008.07.022
6. He Y, Wan S, Hua Y, Keep RF, Xi G (2008) Autophagy after experimental intracerebral hemorrhage. J Cereb Blood Flow Metab 28(5):897–905
7. Cordaro M, Impellizzeri D, Paterniti I, Bruschetta G, Siracusa R, De Stefano D, Cuzzocrea S, Esposito E (2014) Neuroprotective effects of Co-ultraPEALut on secondary inflammatory process and autophagy involved in traumatic brain injury. J Neurotrauma. doi:10.1089/neu.2014.3460
8. Lai Y, Hickey RW, Chen Y, Bayir H, Sullivan ML, Chu CT, Kochanek PM, Dixon CE et al (2008) Autophagy is increased after traumatic brain injury in mice and is partially inhibited by the
antioxidant gamma-glutamylcysteinyl ethyl ester. J Cereb Blood Flow Metab 28(3):540–550
9. Levine B, Klionsky DJ (2004) Development by self-digestion: mo-lecular mechanisms and biological functions of autophagy. Dev Cell 6(4):463–477
10. Melendez A, Talloczy Z, Seaman M, Eskelinen EL, Hall DH, Levine B (2003) Autophagy genes are essential for dauer develop-ment and life-span extension in C. elegans. Science 301(5638): 1387–1391. doi:10.1126/science.1087782
11. Gozuacik D, Kimchi A (2004) Autophagy as a cell death and tumor suppressor mechanism. Oncogene 23(16):2891–2906. doi:10.1038/ sj.onc.1207521
12. Larsen KE, Sulzer D (2002) Autophagy in neurons: a review. Histol Histopathol 17(3):897–908
13. Park JM, Tougeron D, Huang S, Okamoto K, Sinicrope FA (2014) Beclin 1 and UVRAG confer protection from radiation-induced DNA damage and maintain centrosome stability in colorectal can-cer cells. PLoS One 9(6):e100819. doi:10.1371/journal.pone. 0100819
14. Ferraro E, Cecconi F (2007) Autophagic and apoptotic response to stress signals in mammalian cells. Arch Biochem Biophys 462(2): 210–219
15. Chen HC, Fong TH, Lee AW, Chiu WT (2012) Autophagy is acti-vated in injured neurons and inhibited by methylprednisolone after experimental spinal cord injury. Spine (Phila Pa 1976) 37(6):470– 475. doi:10.1097/BRS.0b013e318221e859
16. Kanno H, Ozawa H, Sekiguchi A, Yamaya S, Itoi E (2011) Induction of autophagy and autophagic cell death in damaged neu-ral tissue after acute spinal cord injury in mice. Spine (Phila Pa 1 9 7 6 ) 3 6 ( 2 2 ) : E 1 4 2 7– E 1 4 3 4 . d o i :1 0 . 1 0 9 7 / B R S . 0b013e3182028c3a
17. Liu G, Detloff MR, Miller KN, Santi L, Houle JD (2012) Exercise modulates microRNAs that affect the PTEN/mTOR pathway in rats after spinal cord injury. Exp Neurol 233(1):447–456. doi:10.1016/j. expneurol.2011.11.018
18. Paterniti I, Impellizzeri D, Di Paola R, Navarra M, Cuzzocrea S, Esposito E (2013) A new co-ultramicronized composite including palmitoylethanolamide and luteolin to prevent neuroinflammation in spinal cord injury. J Neuroinflammation 10:91. doi:10.1186/ 1742-2094-10-91
19. Esposito E, Paterniti I, Mazzon E, Genovese T, Di Paola R, Galuppo M, Cuzzocrea S (2011) Effects of palmitoylethanolamide on release of mast cell peptidases and neurotrophic factors after spinal cord injury. Brain Behav Immun 25(6):1099–1112. doi:10. 1016/j.bbi.2011.02.006
20. Bethea JR, Castro M, Keane RW, Lee TT, Dietrich WD, Yezierski RP (1998) Traumatic spinal cord injury induces nuclear factor-kappaB activation. J Neurosci 18(9):3251–3260
21. Kwon BK, Tetzlaff W, Grauer JN, Beiner J, Vaccaro AR (2004) Pathophysiology and pharmacologic treatment of acute spinal cord injury. Spine J 4(4):451–464. doi:10.1016/j.spinee.2003.07.007
22. Esposito E, Cuzzocrea S (2011) Anti-TNF therapy in the injured spinal cord. Trends Pharmacol Sci 32(2):107–115. doi:10.1016/j. tips.2010.11.009
23. Zhang HY, Wang ZG, Wu FZ, Kong XX, Yang J, Lin BB, Zhu SP, Lin L et al (2013) Regulation of autophagy and ubiquitinated pro-tein accumulation by bFGF promotes functional recovery and neu-ral protection in a rat model of spinal cord injury. Mol Neurobiol 48(3):452–464. doi:10.1007/s12035-013-8432-8
24. Mizushima N (2007) Autophagy: process and function. Genes Dev 21(22):2861–2873
25. Baehrecke EH (2005) Autophagy: dual roles in life and death? Nat Rev Mol Cell Biol 6(6):505–510
26. Debnath J, Baehrecke EH, Kroemer G (2005) Does autophagy contribute to cell death? Autophagy 1(2):66–74
27. Reef S, Zalckvar E, Shifman O, Bialik S, Sabanay H, Oren M, Kimchi A (2006) A short mitochondrial form of p19ARF induces autophagy and caspase-independent cell death. Mol Cell 22(4): 463–475. doi:10.1016/j.molcel.2006.04.014
28. Mathew R, Karantza-Wadsworth V, White E (2007) Role of au-tophagy in cancer. Nat Rev Cancer 7(12):961–967. doi:10.1038/ nrc2254
29. Zhong Y, Wang QJ, Li X, Yan Y, Backer JM, Chait BT, Heintz N, Yue Z (2009) Distinct regulation of autophagic activity by Atg14L and Rubicon associated with Beclin 1-phosphatidylinositol-3-kinase complex. Nat Cell Biol 11(4):468–476. doi:10.1038/ncb1854
30. Park JM, Huang S, Wu TT, Foster NR, Sinicrope FA (2013) Prognostic impact of Beclin 1, p62/sequestosome 1 and LC3
protein expression in colon carcinomas from patients receiving 5-fluorouracil as adjuvant chemotherapy. Cancer Biol Ther 14(2): 100–107. doi:10.4161/cbt.22954
31. Tanida I, Ueno T, Kominami E (2008) LC3 and Autophagy. Methods Mol Biol 445:77–88. doi:10.1007/978-1-59745-157-4_4
32. Bjorkoy G, Lamark T, Pankiv S, Overvatn A, Brech A, Johansen T (2009) Monitoring autophagic degradation of p62/SQSTM1. Methods Enzymol 452:181-197. doi:10.1016/S0076-6879(08) 03612-4S0076-6879(08)03612-4
33. Pyronnet S, Sonenberg N (2001) Cell-cycle-dependent translational control. Curr Opin Genet Dev 11(1):13–18
34. Schmelzle T, Hall MN (2000) TOR, a central controller of cell growth. Cell 103(2):253–262