1. Introduzione
Il presente lavoro di tesi analizza la composizione delle faune a Teleostei dei più importanti giacimenti fossiliferi del Cretaceo medio, del Cretaceo superiore e della porzione inferiore Terziario. Lo scopo di questo studio è quello di esaminare il percorso evolutivo del più diversificato taxon di Teleostei attuali, quello degli Acanthomorpha.
Le informazioni ricavate dalla stratigrafia, dalla datazione, dal paleoambiente, dalla paleontologia sistematica e dalla percentuale relativa dei generi acantomorfi (sul totale dei generi Teleostei) per ciascuna delle località fossilifere consentono di contestualizzare meglio, nello spazio e nel tempo, i vari eventi verificatisi nel corso della storia evolutiva degli Acanthomorpha durante l’ultima fase del Mesozoico.
La storia degli Acanthomorpha è stata segnata dalla loro enorme radiazione evolutiva manifestatasi alla base del Cenozoico. Questo episodio ha prodotto la più grande diversificazione della storia dei Vertebrati: basta pensare che attualmente più del 60% delle specie di Teleostei (che rappresentano da soli più della metà delle specie di Vertebrati esistenti) appartengono al taxon Acanthomorpha. I dati elaborati in questo studio, e confrontati con le recenti ipotesi, consentono di tracciare un quadro evolutivo molto interessante in cui gli acantomorfi, fin dai primi momenti della loro storia (iniziata verosimilmente nel Cretaceo medio) manifestarono già gli adattamenti morfologico – funzionali che hanno determinato il loro successo a partire dalle prime fasi del Cenozoico.
1. 1. Caratterizzazione sistematica degli Acanthomorpha
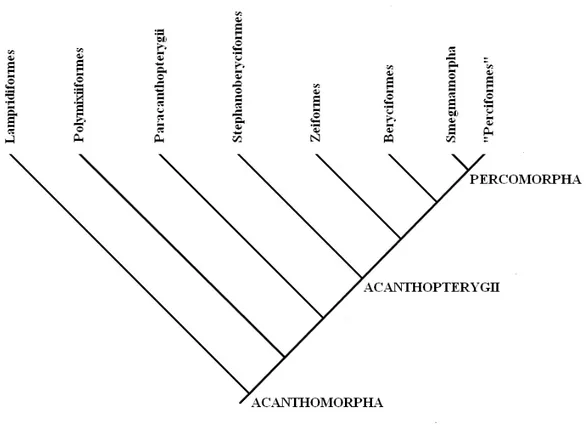
Secondo la definizione presentata nell’articolo “Percomorph phylogeny: a survey of acanthomorphs and a new proposal”, Johnson & Patterson (1993) affermano che il taxon Acanthomorpha debba includere tutti i teleostei che possiedono una pinna dorsale ed anale munita di spine; il carattere monofiletico di questo grande gruppo è fondato su questa sinapomorfia. Attraverso i loro studi sulle relazioni filetiche degli acantomorfi, i due autori identificano sette taxa terminali monofiletici (figura 1): Lampridiformes, Polymixiiformes, Paracanthopterygii, Stephanoberyciformes, Zeiformes, Beryciformes e Smegmamorpha. L’unico gruppo che risulta veramente problematico è il sister – clade degli Smegmamorpha: questo ramo filetico include gli ordini Scorpaeniformes, Perciformes, Tetraodontiformes e Pleuronectiformes.
L’ordine dei Lampridiformes rappresenta lo stem – clade degli Acathomorpha: è composto da 50 specie viventi di predatori meso e batipelagici; la taglia di queste specie è molto variabile, dai 30 cm fino ai 12 m del re delle aringhe (Regalecus glesne).
Figura 1. Cladogramma del taxon Acanthomorpha proposto da Johnson & Patterson (1993).
Polymixia rappresenta l’unico genere vivente dell’ordine dei Polymixiiformes; questi pesci popolano attualmente i fondali di elevata profondità (oltre gli 800 m) e possiedono un paio di caratteristici barbigli prodotti da una modificazione delle ossa dell’apparato ioideo.
Il clade dei Paracanthopterygii è composto da cinque ordini: Percopsiformes, Batrachoidiformes, Lophiiformes, Gadiformes e Ophidiiformes. Johnson & Patterson (1993), pur non mettendo in discussione lo status monofiletico dei Paracanthopterygii, ammettono che le evidenze a sostegno di questa posizione sono molto scarse.
Quello dei Percopsiformes è un piccolo ordine formato da 9 specie di piccoli pesci di taglia compresa tra i 5 ed i 20 cm che popolano le acque continentali del Nord America.
I pesci rospo costituiscono l’ordine dei Batrachoidiformes (la parola greca “batrakhos” significa appunto rospo); sono privi di scaglie, possiedono una colorazione mimetica, mentre la loro testa e la loro bocca sono larghe; la taglia varia dai 5 ai 60 cm. I batracoidiformi sono tutte specie predatrici bentoniche, popolano substrati sabbiosi e rocciosi e tendono agguati alle loro prede restando immobili sul substrato; quasi tutti sono marini.
Le rane pescatrici sono incluse nell’ordine dei Lophiiformes; il loro nome comune è riferito alla forma del corpo ed al loro caratteristico modo di procacciarsi il cibo, facendo compiere dei movimenti ad un’escrescenza carnosa (detta illicio o esca), posizionata all’apice della prima spina della pinna dorsale shiftata anteriormente sul cranio, che serve ad attrarre le prede. Quest’ordine comprende specie pelagiche (compresse lateralmente) e bentoniche (compresse dorso – ventralmente) che popolano i mari di tutto il mondo.
Il piccolo ordine dei Gadiformes comprende i merluzzi e le specie affini. I gadiformi sono distribuiti nelle acque marine di tutto il mondo; le loro dimensioni variano da 10 cm a 2 m ed in seguito ad una riduzione secondaria, le loro pinne non presentano spine.
L’ordine degli Ophidiiformes è formato da acantomorfi pelagici, bentonici e parassiti che popolano svariati tipi di ambienti marini, anche quelli più estremi: Abyssobrotula galatheae, una specie abissale, è stata trovata a 8.370 m, nella fossa di Porto Rico. Molte specie di quest’ordine popolano le acque tropicali e subtropicali in prossimità dei reefs corallini, altre vivono nei mari di entrambi i Circoli Polari; infine, poche specie sono distribuite nelle acque continentali. Il corpo degli ofidiiformi è affusolato, la loro testa è piccola e spesso sono privi di scaglie; le loro lunghe pinne dorsale ed anale sono tipicamente unite a quella caudale, mentre la loro taglia varia dai 5 cm ai 2 m. L’ordine degli Stephanoberyciformes annovera 45 specie marine attuali che vivono ad elevate profondità. Un altro piccolo ordine di pesci marini è quello degli Zeiformes, che include circa 40 specie bentoniche che prediligono i substrati morbidi (sabbiosi e fangosi); gli zeiformi sono distribuiti in tutti gli oceani e vivono generalmente tra i 50 e gli 800 m di profondità. Nel loro studio, Johnson & Patterson (1993) accettano gli Zeiformes come gruppo monofiletico escludendo la famiglia dei Caproidae, la quale sembra possedere dei caratteri affini ai Perciformes.
L’ordine dei Beryciformes comprende circa 200 specie attuali, molte delle quali popolano le acque marine profonde compiendo delle migrazioni notturne verso la superficie. Le specie della famiglia Holocentridae popolano le acque tropicali di tutti gli oceani in prossimità dei reefs corallini; questa famiglia raggiunge il suo picco di diversità specifica in corrispondenza dell’area Indo – Pacifica. Il clade degli Smegmamorpha è stato proposto proprio da Johnson & Patterson (1993); questo raggruppamento comprende i taxa dei Synbranchioidei, Mastacembaloidei, Elassomatidae, Gasterosteiformes (spinarelli, pesci trombetta, cavallucci marini…), Mugilomorpha (muggini) ed Atherinomorpha. Il nome di questo taxon deriva dalla parola greca “smegma”, che significa detergere, ed è stata scelta per sottolineare l’intento dei due autori di voler chiarire l’inquadramento sistematico degli acantomorfi più evoluti. Secondo Johnson & Patterson (1993) gli Smegmamorpha possiedono un’unica sinapomorfia: la prima coppia di epineurali si sviluppa alla base delle
parapofisi della prima vertebra (mentre nella condizione plesiomorfica i primi epineurali si inseriscono alla base della prima spina neurale).
Nel definire la sistematica dei Percomorpha, Johnson & Patterson (1993) premettono che la composizione originale di questo taxon, introdotto per la prima volta da Rosen (1973), non ha alcun fondamento naturale e definibile in senso cladistico. Secondo i due autori, i Percomorpha possono costituire un gruppo naturale attraverso la separazione in due sister – taxa: il primo è quello degli Smegmamorpha; il secondo annovera circa 9.300 specie attuali, ripartite negli ordini dei Perciformes, Scorpaeniformes, Pleuronectiformes e Tetraodontiformes.
Con oltre 7.000 specie viventi, quello dei Perciformes rappresenta il più grande ordine di vertebrati. I perciformi sono diffusi in ogni tipo di ambiente acquatico ed hanno raggiunto le taglie più disparate (dai 7 mm di Schindleria brevipinguis agli oltre 5 m delle specie appartenenti al genere Makaira). Tuttavia, l’ordine dei Perciformes è molto problematico: nelle classificazioni tradizionali, come quelle di Greenwood et al. (1966) e di Nelson (1984), questo taxon è stato sempre collocato tra i cosiddetti ordini “pre – perciformi” (come gli Scorpaeniformes) e gli ordini più derivati, definiti per questo “post – perciformi” (Pleuronectiformes e Tetraodontiformes); i Perciformes, quindi, sarebbero un taxon parafiletico, ma questa posizione non può essere accettata in termini cladistici. Johnson & Patterson (1993), pur non esprimendo alcun giudizio definitivo sulla questione, sottolineano l’importanza di considerare i Perciformes ed i taxon a loro più prossimi (Scorpaeniformes, Pleuronectiformes e Tetraodontiformes) come sister – clade degli Smegmamorpha.
L’ordine dei Tetraodontiformes annovera circa 360 specie, la maggioranza delle quali popola i mari tropicali dei reefs corallini o in prossimità di essi. I tetraodontiformi popolano le acque marine tropicali e temperate di tutto il mondo; alcune specie vivono nelle acque continentali dell’Africa, Sud America ed Asia sud orientale. Le loro dimensioni sono molto variabili, dai 2 cm di lunghezza del genere Rudarius agli oltre 3 m (e 2 tonnellate di peso) del genere Mola.
L’ordine degli Scorpaeniformes è composto da pesci marini predatori che popolano generalmente i fondali marini poco profondi cibandosi di crostacei e piccoli pesci. Gli scorpeniformi possiedono
una testa spinosa, mentre le pinne caudale e pettorali hanno una tipica forma tondeggiante; la maggior parte di essi non raggiunge i 30 cm di lunghezza.
I “pesci piatti” (sogliole, rombi, halibut…) costituiscono l’ordine dei Pleuronectiformes. Le oltre 400 specie di pleuronettiformi popolano i substrati marini sabbiosi e rocciosi, e molte esse possiedono delle colorazioni mimetiche. Quest’ordine è noto anche col nome di Heterosomata, a causa dell’asimmetria corporea che questi pesci sviluppano nel corso dell’ontogenesi: le forme giovanili presentano una normale simmetria bilaterale; successivamente un occhio (di solito il
sinistro) migra sulla cima della testa andando a situarsi sull’altro lato, al di sopra del secondo occhio; il cranio diventa notevolmente compresso e subisce deformazioni permanenti; queste modificazioni, insieme ad altre, consentono a questi pesci di poggiarsi o infossarsi sul fondale in attesa di una preda.
Attualmente, lo studio di Johnson & Patterson (1993) costituisce un punto di riferimento essenziale per comprendere le relazioni tra i cladi acantomorfi più importanti (figura 1); tuttavia, mentre i rapporti filogenetici tra i gruppi più basali appaiono chiari (ad eccezione dei Paracanthopterygii), non si può dire lo stesso per i Percomorpha, dove un numero notevole di taxa non ha una collocazione sistematica precisa.
2. Analisi descrittiva dei giacimenti a pesci del Cretaceo medio e superiore
2. 1. I depositi di acantomorfi nel Cretaceo medio e superiore
In questa sezione sono state raccolte le informazioni relative alla stratigrafia, alla datazione, al paleoambiente, alla paleontologia sistematica ed alla percentuale relativa dei generi acantomorfi (sul totale dei generi Teleostei) di ciascuna delle località fossilifere del Cretaceo medio e superiore esaminate in questa tesi. I seguenti paragrafi descrittivi sono disposti in sequenza temporale, partendo dalle località fossilifere più antiche. L’elenco delle ittiofaune a Teleostei di tutti i depositi mesozoici esaminati in questo lavoro di tesi sono consultabili nella Tavola 1 (pag. 26 ). I grafici che illustrano la percentuale di abbondanza relativa dei generi acantomorfi per ciascuna ittiofauna a Teleostei esaminata in questo lavoro di tesi sono stati raccolti nella Tavola 2 (pag. 30).
2. 1. 1. Hakel e Hadjoula (Libano)
Stratigrafia e datazione. Le ittiofaune cenomaniane più famose del Libano sono quelle di Hakel e Hadjoula: la prima è nota fin dai tempi delle crociate, mentre le prime documentazioni riguardanti Hadjoula risalgono all’ultimo decennio del XIX secolo. Le due località, molto vicine tra loro, si trovano ad una cinquantina di km a Nord Est di Beirut (Basson & Edgell, 1971).
I depositi carbonatici cenomaniani del Libano (compresi quelli dei siti di Hakel, Hadjoula e Nammoura) fanno parte di un’unica unità geologica, i “Calcari di Sannine”, spessa fino a 650 m ed estesa dalla costa mediterranea alla dorsale dei Monti del Libano; l’età cenomaniana dei “Calcari di Sannine” è stata stabilita attraverso il riconoscimento di un foraminifero bentonico (Pseudorhapydiolina laurinensis) contenuto nelle rocce di questa unità litologica (de Castro, 1965).
Gli strati a pesci di Hakel e Hadjoula, inoltre, contengono entrambi il foraminifero Orbitolina concava, che ha consentito datare i due depositi al Cenomaniano inferiore (Hükel, 1970).
Paleoecologia. I depositi fossiliferi di Hakel e Hadjoula sono entrambi marini, di acque poco profonde. La frazione dominante della fauna dei due depositi è quella ittica (con pesci ossei e cartilaginei); scarsi, invece, sono i fossili di organismi terrestri, sia animali che vegetali, il che implica una certa lontananza dell’ambiente di deposizione dalla linea costa (Forey et al., 2003). Il numero di pesci fossili rinvenuto è molto elevato: spesso, questi reperti si concentrano in lastre calcaree; tale disposizione si è realizzata in conseguenza ad eventi di mortalità di massa legati a fioriture fitoplanctoniche. Molto probabilmente, l’ambiente di deposizione di entrambe le località ha subito ripetuti movimenti tettonici che hanno confinato i pesci in vere e proprie “conche – trappola” mortali (Hükel, 1970).
2. 1. 2. Nammoura (Libano)
Stratigrafia e datazione. Il deposito di Nammoura si trova nei pressi dell’omonimo villaggio, nella valle di Al Ghabour, in prossimità della costa a nord di Beirut. Durante il XX secolo sono state aperte due cave fossilifere in corrispondenza di un affioramento calcareo lungo circa 1 km. La roccia di questo affioramento è un tipico calcare litografico di micrite chiara, laminata e ricca di lenti di selce (Forey et al., 2003); lo stato di conservazione dei fossili è eccezionale. Gli strati a pesci di Nammoura, spessi 30 m, consistono in calcari litografici tabulari, di colore giallo o marrone chiaro, a laminazione piano parallela (Dalla Vecchia et al., 2002) ed apparentemente privi di microfossili. Tale assenza ha reso complicata l’analisi di datazione del deposito di Nammoura. Solo recentemente, uno studio sulla stratigrafia dell’intero affioramento compiuto da Dalla Vecchia et al.
(2002) ha evidenziato la presenza di una ricca microfauna di foraminiferi del Cenomaniano superiore nei calcari che sovrastano immediatamente gli strati a pesci;
perciò, l’ittiofauna di Nammoura è stata datata al Cenomaniano medio. Paleoecologia. I pesci (ossei e cartilaginei) costituiscono la grande maggioranza dei reperti di
Nammoura; la fauna è completata da alcuni fossili di crostacei, echinoidi, tartarughe, serpenti, un dolicosauro (uno squamato marino) ed un dinosauro teropode (Dalla Vecchia et al., 2002). Anche i fossili di piante (Krassilov & Bacchia, 2000; Dilcher & Basson, 1990) sono piuttosto abbondanti, con una netta prevalenza di angiosperme: la loro presenza nel record di Nammoura, insieme a quella dei fossili animali terrestri, dimostra che l’ambiente di deposizione doveva essere in prossimità della costa. In conclusione, il paleoambiente di Nammoura era di tipo marino, di acque superficiali, particolarmente vicino alla terraferma; tuttavia, dato che nel Cenomaniano la linea di
costa del continente più vicino a Nammoura (il Gondwana) correva a centinaia di km più a sud (Philip et al., 1993), è plausibile pensare che questo deposito rappresenti il residuo di una formazione carbonatica di un’isola nel bacino della Tetide (Forey et al., 2003).
2. 1. 3. Jebel Tselfat (Marocco)
Stratigrafia e datazione. Il deposito marino di Jebel Tselfat affiora in due località (An el Kerma e
Sigda) a Nord Est di Fes, nel Marocco settentrionale. Gli strati fossiliferi sono costituiti da un argillite nerastra. I pesci si presentano parzialmente disarticolati: questo indica che i loro resti hanno subito un’azione di trasporto all’interno dell’ambiente di sedimentazione
(Forey et al., 2003). La datazione di Jebel Tselfat è stata discussa da Cavin & Dutheil (1999)
sulla base della comparazione dei suoi ittioliti con quelli rinvenuti recentemente nel giacimento fossilifero marocchino di Daura; da questo esame, i due studiosi hanno stabilito
che gli ittioliti di Jebel Tselfat risalgano al Cenomaniano superiore. Paleoecologia. Le argilliti si sono originate in un ambiente marino di moderata profondità e lontano
dalla costa (Arambourg, 1954). L’ittiofauna di Jebel Tselfat è composta da pochi picnodonti e clupeomorfi (abitanti tipici delle acque poco profonde), mentre è ricca in ittioliti di elopiformi e polimixiiformi, due taxa che attualmente includono forme di ambiente pelagico e di acque profonde (Forey et al., 2003).
2. 1. 4. Chalk Group (Inghilterra meridionale)
Stratigrafia e datazione. Il “Chalk Group” è un’unità litostratigrafica distribuita soprattutto nella parte nord occidentale dell’Europa; tale unità è caratterizzata da spessi accumuli di calcare di origine marina depositatosi durante il Cretaceo superiore (dal Cenomaniano al Campaniano). I calcari del “Chalk Group” sono costituiti da pareti calcitiche di alghe unicellulari (Chrysophyceae, Coccolithophorales). In Inghilterra, le formazioni che compongono il “Chalk Group” sono tre: Upper Chalk (la più recente e superficiale), Middle Chalk e Lower Chalk (la più profonda ed antica). I famosi e spettacolari affioramenti del “Chalk Group” inglese, come le scogliere di Dover o quelle dell’isola di Wight, si concentrano nella parte meridionale del paese. Lo spessore totale dell’unità litostratigrafica supera i 300 m. La formazione inferiore è sicuramente quella più ricca di fossili. Gli strati della sua base sono costituiti da marne cloritiche e glauconitiche dal caratteristico colore verde; procedendo verso l’alto, la frazione argillosa si riduce e gli strati marnosi lasciano il posto a
Il record fossile della formazione Lower Chalk è caratterizzato da una consistente malacofauna (ammoniti, belemniti, bivalvi ostreidi ed inoceramidi); sono stati rinvenuti anche diversi esemplari di echinodermi (asteroidei ed echinoidei) e briozoi; infine, l’ittiofauna è rappresentata da una quarantina di generi di pesci ossei e cartilaginei. Gli unici generi di pesci dell’English Chalk che hanno avuto un preciso inquadramento stratigrafico sono proprio quelli della formazione Lower Chalk, datati al Cenomaniano superiore con il metodo biostratigrafico: i livelli a pesci corrispondono alle biozone dell’ammonite
Schloenbachia varians e dell’asteroideo Holoaster subglobosus (Forey et al., 2003). Paleoecologia. Le formazioni inglesi del Cretaceo superiore costituiscono i resti di un grande
sistema di piattaforme carbonatiche marine che si svilupparono nella fascia paleoclimatica subtropicale – temperato calda dell’Europa nel periodo finale del Mesozoico. Il fondale della piattaforma inglese doveva trovarsi a circa 200 m di profondità (Scholle et al., 1983). L’ittiofauna della formazione Lower Chalk rispecchia ampiamente la natura del suo ambiente di deposizione: gran parte dei generi teleostei qui rinvenuti appartengono ad ordini tipicamente pelagici (Ichthyodectiformes, Elopiformes, Aulopiformes, Polymixiiformes); questa predominanza si riscontra anche analizzando il record fossile dei condroitti.
2. 1. 5. Komen (Slovenia)
Stratigrafia e datazione. Le rocce che affiorano nei dintorni di Komen costituiscono il cuore del “Trieste – Komen plateau”, un’unità geologica di piattaforma carbonatica di età cretacea spessa oltre 2000 m (Palci et al., 2007). Gli affioramenti di Komen sono formati da rocce calcaree grigio scure, bituminose e finemente stratificate (Jurkovšek et al., 1996; Cavin et al., 2000). La datazione cenomaniana degli strati fossiliferi di Komen è stata effettuata attraverso l’identificazione di un fossile guida, il foraminifero Broeckina (Pastrikella) balcanica (Cavin et al., 2000). Tra i fossili di questi depositi, i pesci costituiscono la frazione preponderante, sebbene anche i resti di rettili (Nopcsa, 1903; Calligaris, 1988) e di piante (Dobruskina et al., 1999) siano abbastanza comuni ; i
fossili di invertebrati (rudiste, ammoniti, crostacei, asteroidei) sono molto rari (Palci et al., 2007). Paleoecologia. La composizione dell’ittiofauna di Komen testimonia il carattere marino tropicale (o
sub – tropicale) dell’ambiente in cui questi pesci vivevano. Inoltre, la presenza di un numero consistente di ritrovamenti attribuiti a rettili semi – acquatici e piante ha permesso di definire meglio l’ambiente di deposizione, caratterizzato da acque poco profonde e piuttosto vicino all’antica linea costiera (Palci et al., 2007). Questo ambiente ricco di prede ha attratto i pesci pelagici verso la costa, come dimostrano i ritrovamenti di alcuni ictiodectiformi. Resta da chiarire la
presenza di alcuni fossili di taxa tipicamente batipelagici (come gli aulopiformi ed i mictofiformi): è probabile che questi gruppi di pesci popolassero l’ambiente neritico e / o epipelagico durante il
Cretaceo superiore, per poi migrare verso le grandi profondità alla fine di questo periodo. In conclusione, gli ittioliti di Komen costituiscono la frazione dominante di una fauna marina del
Cenomaniano che popolò le acque di una piccola piattaforma intraoceanica settentrionale del bacino della Tetide (Palci et al.,2007).
2. 1. 6. Cinto Euganeo (Padova, Italia settentrionale)
Stratigrafia e datazione. Il giacimento di Cinto Euganeo si trova ai piedi del Monte Cinto, dove i lavori di sbancamento di una grande cava di cemento (Cava Bomba) hanno portato alla luce una serie di strati di colore scuro. Questi strati appartengono ad una lente dello spessore massimo di 4 m, estesa per circa 40 m ed interposta a livelli di calcari lastriformi di colore grigio chiaro e rosato. Il livello fossilifero è costituito da un’argillite bituminosa nerastra, finemente stratificata, con lamine ricche di marcasite e pirite, entrambi minerali di solfuro di Ferro (Sorbini, 1976). Gli strati fossiliferi sono privi di microfossili stratigraficamente significativi; tuttavia, immediatamente alla base della lente a pesci affiorano dei calcari che contengono foraminiferi planctonici della zona a Rotalipora cushmani, del Cenomaniano superiore, mentre i calcari al tetto della lente contengono un’associazione di foraminiferi della zona a Hedbergella lehmanni, del Turoniano inferiore. Pertanto il giacimento di Cinto Euganeo è stato datato al limite Cenomaniano – Turoniano (Sorbini, 1976).
Paleoecologia. Da un punto di vista ecologico, l’esame dei generi trovati a Cinto Euganeo dimostra il carattere esclusivamente marino dell’ittiofauna. Come per altri giacimenti cretacei, nel deposito di Cinto Euganeo sono presenti diversi generi appartenenti ad ordini (Myctophiformes, Aulopiformes, Polymixiiformes…) i cui rappresentanti attuali vivono a grande profondità. Tuttavia, l’assenza di
fotofori ed il mancato riscontro di una regressione nell’ossificazione sui reperti, al pari di quelli di Floresta e di Jebel Tselfat, rende probabile l’ipotesi che l’adattamento
alla vita nelle acque profonde sia stato acquisito in tempi successivi (Sorbini, 1976). Dall’esame complessivo dei generi che costituiscono l’ittiofauna di Cinto Euganeo si può affermare che essa ha principalmente carattere pelagico, inserita nel contesto tropicale del bacino tetideo e che la presenza di forme durofaghe (soprattutto tra i generi condroitti) indica una non eccessiva profondità del mare. Da un punto di vista biogeografico, la fauna di Cinto Euganeo presenta le maggiori affinità con quelle di Jebel Tselfat e di Floresta, dato il numero elevato di generi condivisi (Sorbini, 1976).
2. 1. 7. Floresta (Messina, Sicilia nord orientale)
Stratigrafia e datazione. Gli affioramenti ittiolitici cretacei della catena montuosa nebrodico – peloritana appartengono all’unità geologica delle cosiddette “Argille Scagliose Antisicilidi”, comprese originariamente nel “Complesso Sicilide”, dal quale poi si sono separate attraverso un particolare meccanismo di retrocolamento gravitativo (Ogniben, 1969; Guerrera & Wezel, 1974; Lentini et al., 1987). Il “Complesso Sicilide” rappresenta ciò che resta dallo scollamento del fondo oceanico dell’antica Tetide sotto la pressione imposta dall’orogenesi appenninico – maghrebide, verificatasi durante il Terziario a causa della collisione tra la placca euroasiatica e quella africana (Lentini et al., 2000).
Le “Argille Scagliose Antisicilidi” di Floresta e delle altre località fossilifere distribuite nella provincia di Messina sono rappresentate da una regolare successione dal basso verso l’alto di strati di argille, argilloscisti, e radiolariti (Leonardi, 1966; Campisi, 1977). I livelli fossiliferi sono costituiti dagli strati bituminosi grigio scuri e neri e dalla parte basale delle soprastanti radiolariti di colore più chiaro, ma le numerose fratture ortogonali ai piani di strato dimostrano le alterne vicende a cui sono andate incontro queste formazioni prima della loro definitiva messa in posto (Rindone, 2008).
La potenza degli strati varia dai 20 m dei livelli di Floresta ai 90 m della successione sedimentaria quasi completa di Malvagna, nella quale gli strati ittiolitici si concentrano nei primi 20 m (Rindone, 2008).
La datazione al limite Cenomaniano – Turoniano dei fossili siciliani è stata fornita dall’analisi delle associazioni a radiolari, nannofossili calcarei e foraminiferi planctonici presenti negli strati a pesci (Bellanca et al., 1998); inoltre, nei medesimi strati è stato individuato il “Livello Bonarelli”, l’espressione sedimentaria nelle successioni italiane di un evento anossico oceanico di scala mondiale verificatosi proprio in corrispondenza di questo limite (Bellanca et al., 1998; Bellanca et al., 2000). Questo livello è stato individuato anche nei depositi di Cinto Euganeo e di Jebel Tselfat. Paleoecologia. L’analisi della composizione specifica dei pesci fossili siciliani ed il tipo di sedimento consentono di stabilire che si tratta di un’ittiofauna composita di mare caldo, con un carattere prevalentemente meso – batipelagico, che si è depositata in un bacino profondo, con scarsa o assente circolazione delle acque, i cui strati inferiori erano totalmente anossici. La conservazione dei fossili è stata possibile grazie all’aumento del tasso di sedimentazione, alla presenza di acque calme e stagnanti, alle condizioni riducenti del fondo asfittico per l’abbondanza di H2S ed alla totale assenza di organismi detritivori e necrofagi, distruttori della sostanza organica (Rindone, 2008).
2. 1. 8. Bohemian Cretaceous Basin (Repubblica Ceca)
Stratigrafia e datazione. Il Bohemian Cretaceous Basin rappresenta un’area paleobiogeografica caratteristica, localizzata in un braccio di mare che metteva in comunicazione la Tetide settentrionale ed il grande sistema di piattaforme carbonatiche boreali della fascia paleoclimatica temperata (Wiese et al., 2004; Ekrt et al., 2008).
Tra le diverse località fossilifere del Bohemian Cretaceous Basin, sicuramente la cava Upolhavy (a circa 70 km a nord est di Praga) è quella che ha portato alla luce i reperti più importanti per numero e qualità. La sezione inferiore della cava presenta degli strati di marne scure; questa composizione omogenea è interrotta, nella parte superiore, da un’alternanza di strati di marne e calcari. I pesci fossili (ossei e cartilaginei) sono stati rinvenuti in associazione a resti di rettili marini (probabilmente plesiosauri); la presenza di un’abbondante fauna di invertebrati (tra cui gasteropodi, bivalvi e ammoniti) ha permesso di datare con precisione il deposito, facendolo risalire al Turoniano superiore (Čech et al., 1996; Ekrt et al., 2001; Wiese et al., 2004).
Paleoecologia. La composizione ittiologica del record fossile del Bohemian Cretaceous Basin presenta una forte somiglianza con l’ittiofauna del Chalk Group inglese. La profondità del Bohemian Cretaceous Basin superava raramente i 100 m: questo spiega la presenza di numerosi fossili di picnodonti (attinopterigi non teleostei durofagi) e dell’acantomorfo Hoplopteryx (Ekrt et al., 2008). I fossili presentano una caratterizzazione ecologica simile a quella di altre faune boreali del Creaceo superiore, nelle quali si nota una sproporzione tra i vertebrati piscivori (sempre abbondanti e diversificati) e quelli che si nutrono di invertebrati o di plancton (Ekrt et al., 2008).
2. 1. 9. Smoky Hill (Kansas, U.S.A.)
Stratigrafia e datazione. L’unità geologica dei “Calcari di Smoky Hill” è inclusa nella formazione Niobrara, che comprende la maggior parte dei depositi tardo cretacei dello stato del Kansas.
I “Calcari di Smoky Hill” sono il prodotto di una sedimentazione marina iniziata nel Coniaciano superiore e protrattasi fino al Campaniano inferiore (87 – 82 milioni di anni fa), quando un grande mare epicontinentale (il Western Interior Sea) divideva longitudinalmente il Nord America mettendo in comunicazione le acque del Circolo Polare Artico con quelle dell’attuale Golfo del Messico (Everhart, 2005).
Lo spessore dei “Calcarei di Smoky Hill” è circa 200 m; queste rocce sono principalmente composte da pareti calcitiche di alghe unicellulari (Chrysophyceae, Coccolithophorales) che popolavano acque limpide. Nei “Calcari di Smoky Hill” si intercalano un centinaio di strati argillosi
rosso ruggine di origine vulcanica. Questi depositi sono stati utilizzati da Hatting (1982) per descrivere e datare l’unità geologica in questione; inoltre, molte specie di vertebrati ed invertebrati marini sono stati utilizzati come fossili guida per compiere una datazione più accurata di alcuni orizzonti, come quelli che includono gli strati a pesci.
Paleoecologia. Il bacino epicontinentale del Western Interior Sea non superava i 200 m di profondità; in corrispondenza dell’attuale stato del Kansas, i fondali di questo mare erano piatti, fangosi e profondi soltanto alcune decine di metri.
I “Calcari di Smoky Hill” costituiscono una porzione di piattaforma interna del Western Interior Sea ed hanno conservato un grande numero di organismi fossili che componevano la fauna del tratto centrale di questo mare interno: sono stati rinvenuti bivalvi inoceramidi e rudiste, crinoidi, ammoniti ed altri molluschi cefalopodi, numerosi pesci ossei e cartilaginei, plesiosauri, mosasauri, pterosauri ed uccelli odontognati (Everhart, 2005); la presenza dei fossili di vertebrati terrestri indica che l’ambiente di sedimentazione dei “Calcari di Smoky Hill” doveva essere piuttosto vicino alla linea costiera.
L’ittiofauna di Smoky Hill è composta da predatori piscivori di varia taglia e da forme bentoniche durofaghe. In particolare, il record di teleostei di questo deposito è caratterizzato dalla dominanza di generi piscivori tipicamente pelagici, come quelli inclusi negli ordini Ictiodectiformes, Tselfatiiformes ed Aulopiformes (Shimada & Fielitz, 2006).
In particolare, gli strati a pesci che hanno portato alla luce i fossili di acantomorfi sono dislocati in quattro orizzonti: nel primo, risalente al Coniaciano superiore, è stato rinvenuto il reperto fossile di un bericiforme olocentride; il secondo, del Santoniano inferiore, ha portato alla luce il fossile di un altro bericiforme olocentride, mentre il terzo orizzonte, del Santoniano superiore, è quello che ha fornito il maggior numero di acantomorfi (appartenenti ai generi Kansius, Caproberyx e Trachichthyoides); infine, il quarto ha rivelato la presenza del genere fossile polimixiforme Omosoma. È molto interessante sottolineare che quasi tutti i reperti dei generi acantomorfi di Smoky Hill, alcuni dei quali non ancora descritti, sono stati trovati all’interno delle conchiglie di bivalvi inoceramidi (Stewart, 1990).
2. 1. 10. Nardò (Lecce, Italia sud orientale)
Stratigrafia e datazione. Gli strati calcarei a pesci dei depositi di Nardò appartengono alla porzione superiore dell’unità geologica nota col nome di “Calcari di Melissano”.
Nel loro recentissimo studio cronostratigrafico sulle piattaforme carbonatiche cretacee della penisola salentina, Schlüter et al. (2007) hanno sottoposto svariati campioni delle rocce di
quest’unità ad esami di radiodatazione utilizzando l’isotopo 87 dello Stronzio (87Sr). I risultati di questo studio attribuiscono ai “Calcari di Melissano” un’età media di 82.5 milioni di anni, mentre le successioni più antiche dell’unità risalgono a 85.6 (+/- 0.6) milioni di anni. Dunque, questa recente analisi cronostratigrafica consente di datare il deposito di Nardò al Campaniano inferiore.
Paleoecologia. La piattaforma cretacea pugliese è un tipico esempio di piattaforma carbonatica intraoceanica del bacino della Tetide. La sedimentazione carbonatica iniziò nel tardo Triassico e proseguì per tutto il Mesozoico, producendo un accumulo di rocce spesso circa 6000 m. I depositi cretacei della piattaforma pugliese affiorano lungo la costa adriatica della penisola del Salento, dal promontorio del Gargano fino a Capo Santa Maria di Leuca. Gli affioramenti del Cretaceo superiore sono divisi in tre unità: quella dei “Calcari di Melissano” è la più antica. Gli strati dell’unità di Melissano sono caratterizzati da una scarsa densità di rudiste e dall’assenza di tracce di bioturbazione. Queste informazioni hanno contribuito a ricostruire il paleoambiente di sedimentazione delle rocce carbonatiche di Melissano, uno spazio peritidale separato dal mare aperto per interposizione di una scogliera organogenica, in cui la circolazione delle acque doveva essere molto ridotta. In questo ambiente così circoscritto potrebbero essersi verificati fenomeni di mortalità di massa che hanno così determinato la ricchezza del deposito di Nardò (Sorbini, 1981).
2. 1. 11. Sendenhort e Baumberge (Westfalia, Germania occidentale)
Stratigrafia e datazione. I fossili di Sendenhorst e Baumberge, conosciuti per la loro eccellente qualità da almeno due secoli, sono stati estratti dagli affioramenti carbonatici del Campaniano superiore del bacino di Münster, nella regione della Westfalia (Germania occidentale). Tali depositi sono ripartiti in due membri sovrapposti del bacino: quello superiore di Coesfeld, cui appartengono i fossili di Sendenhorst (Hasemann, 1975; Kaever & Lommerzheim, 1995), e quello inferiore di Baumberge. I depositi sono caratterizzati da un’alternanza di strati argillosi e calcarei di colore grigio e verdastro.
Paleoecologia. Durante il Campaniano superiore, il bacino di Münster costituiva un sistema di comunicazione tra la Tetide e le piattaforme carbonatiche del Dominio Boreale (Dietze, 2008). Le caratteristiche litologiche e biologiche dei depositi di Sendenhorst e Baumberge delineano un ambiente marino profondo (meso – batipelagico) con una lenta circolazione delle acque ed una bassa velocità di sedimentazione (Kaever & Lommerzheim, 1995). L’ittiofauna è composta da taxa neritici (squali), bentonici (anguilliformi e batoidei) e pelagici (aulopiformi, mictofiformi, elopiformi, polimixiformi).
2. 1. 12. Gerusalemme (Israele) e Laveiras (Portogallo)
I dati riguardanti le ittiofaune di Gerusalemme e di Laveiras sono incompleti e tuttora in fase di studio. Gli ittioliti di Gerusalemme sono stati datati al Cenomaniano medio e fanno parte, sulla base delle prime descrizioni paleoambientali, di una più ampia comunità marina di acque poco profonde localizzata in prossimità di un reef. L’ittiofauna portoghese di Laveiras, invece, risale al Cenomaniano medio – superiore ed i suoi depositi fossiliferi si sono anch’essi formati in un ambiente marino di acque poco profonde (Forey et al., 2003).
3. Discussione
4. 1. Gli acantomorfi mesozoici
Tutti i depositi tardo mesozoici esaminati in questa tesi sono marini: dieci di essi sono distribuiti nel bacino della Tetide (Hakel, Hadjoula, Gerusalemme, Jebel Tselfat, Komen, Nammoura, Cinto Euganeo, Floresta, Sahel Alma, Nardò), quattro nel Dominio Boreale (Lower Chalk, Bohemian Cretaceous Basin, Sendenhorst e Baumberge), uno nel Western Interior Sea (Smoky Hill), uno in corrispondenza dell’Atlantico settentrionale (Laveiras).
Inoltre, le analisi sui paleoambienti di sedimentazione rivelano che la grande maggioranza delle località fossilifere si sono formate nella zona neritica: alcune di esse erano particolarmente vicine alla fascia costiera (Komen, Nammoura, Nardò), altre meno (Hakel, Hadjoula, Smoky Hill). Soltanto quattro depositi appartengono ad un ambiente meso – batipelagico (Cinto Euganeo, Floresta, Jebel Tselfat, Sendenhorst e Baumberge).
Dal punto di vista paleoclimatico, le ittiofaune sono quasi tutte distribuite nella fascia paleotropicale; soltanto quattro di esse (Lower Chalk, Bohemian Cretaceous Basin, Smoky Hill, Sendenhorst e Baumberge) sono localizzate nella fascia paleoclimatica temperata boreale.
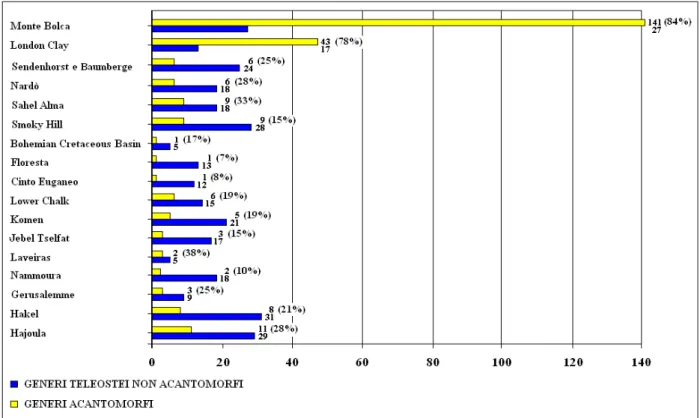
La percentuale dei generi acantomorfi esprime l’abbondanza relativa di questo taxon all’interno di una comunità di teleostei. Dall’analisi delle percentuali ottenute dal record di teleostei di ciascuna località si sono ottenute delle tendenze significative. Dal punto di vista paleoambientale, le ittiofaune di teleostei di zona neritica detengono la percentuale maggiore di generi acantomorfi (che mediamente si attesta intorno al 25%), con un picco positivo a Laveiras (38%) ed uno negativo a Nammoura (10%); nelle ittiofaune meso – batipelagiche del Cretaceo medio e superiore tale percentuale è molto bassa a Cinto Euganeo (8%) e Floresta (7%), datate entrambe al limite Cenomaniano – Turoniano, mentre l’abbondanza relativa di acantomorfi nell’ittiofauna di
Sendenhorst e Baumberge, del Campaniano superiore, raggiunge una percentuale considerevole (25%). Esaminando tale abbondanza relativa dal punto di vista paleoclimatico, ci si accorge che i depositi neritici della fascia paleotropicale presentano generalmente le percentuali più elevate di acantomorfi, mentre nelle località fossilifere neritiche distribuite nella fascia paleoclimatica temperata (Lower Chalk, Bohemian Cretaceous Basin, Smoky Hill) si rilevano delle percentuali minori (figura 2). Le percentuali di abbondanza relativa di acantomorfi sono piuttosto contenute a Floresta e Cinto Euganeo, dove si osservano percentuali inferiori al 10%. L’ittiofauna di Sahel Alma è formata per un terzo da acantomorfi, mentre la percentuale elevata di Laveiras è poco attendibile a causa dell’incompletezza descrittiva della sua ittiofauna.
Figura 2. Diagramma di distribuzione dei generi teleostei delle ittiofaune esaminate in questa tesi; le percentuali tra parentesi esprimono l’abbondanza relativa dei generi acantomorfi.
Da quanto osservato è possibile rilevare come la grande differenza che distingue gli acantomorfi mesozoici dai loro discendenti cenozoici dipenda dal ruolo assunto da questi pesci all’interno delle rispettive comunità ittiche (figura 2): gli acantomorfi terziari ed attuali sono gli assoluti dominatori del panorama ittico (basta pensare che delle 23.600 specie attuali di teleostei, che da soli rappresentano il 90% degli attinopterigi esistenti, 14.600 sono acantomorfe); mentre gli acantomorfi mesozoici costituivano una porzione sempre minoritaria, talvolta marginale, delle ittiofaune.
3. 2. Origine degli acantomorfi
Allo stato attuale delle conoscenze, il taxon Acanthomorpha compare per la prima volta nel record fossile con il genere marino nord americano Muhichthys, risalente al limite Albiano – Cenomaniano. I quattro esemplari di Muhichthys rinvenuti nella cava di Muhi, nello stato federale di Hidalgo in Messico, sono stati descritti da Gonzalez – Rodriguez & Fielitz (2008). Tuttavia, il genere Muhichthys è stato subito relegato tra gli Acanthomorpha incertae sedis, anche se Gonzalez – Rodriguez & Fielitz (2008) hanno identificato dei caratteri che Muhichthys sembra condividere con i polimixiformi ed i bericiformi. La comparsa del Muhichthys precede quella dei generi acantomorfi rinvenuti nelle due località libanesi del Cenomaniano inferiore, Hakel ed Hadjoula. Tuttavia, se si escludono gli acantomorfi di incerta collocazione sistematica, molti dei generi che compongono queste due ittiofaune non sono stati classificati, come ci si potrebbe aspettare, negli ordini basali del taxon Acanthomorpha. Nelle ittiofaune di Hakel ed Hadjoula ci sono diversi generi appartenenti ai Beryciformes, Zeiformes e Tetraodontiformes, tutti ordini che sono inclusi negli Acanthopterygii; inoltre, i Tetraodontiformes fanno parte dei Percomorpha, il crown – group degli Acanthomorpha.
Indubbiamente, la questione dell’origine degli acantomorfi è estremamente complessa; inoltre, l’incompletezza del record paleontologico e il quadro sistematico incerto non contribuiscono a fornire le informazioni necessarie per risolvere questo problema.
Ciò nonostante, l’insieme dei dati sugli acantomorfi mesozoici che finora sono stati raccolti hanno consentito di definire un quadro evolutivo generale per questo grande taxon di teleostei.
Gli acantomorfi più arcaici compaiono nell’ambiente marino. Le evidenze a sostegno di quest’ipotesi sono consistenti: sicuramente il genere Muhichthys popolava un ambiente marino, così come la totalità dei generi acantomorfi cenomaniani. Inoltre, la totalità dei generi più antichi del taxon Acanthomorpha è stata rinvenuta in depositi neritici paleotropicali. Infatti, tutti i generi acantomorfi descritti nell’ittiofauna del Lower Chalk, l’unica località cenomaniana di paleoclima temperato, erano già presenti nei depositi a pesci della Tetide nel Cenomaniano medio ed inferiore. I più antichi acantomorfi mesozoici sono comparsi nel Cretaceo medio, in un momento in cui si stavano verificando dei grandi cambiamenti su scala globale, innescati da un’impennata di attività crostale oceanica e / o di vulcanismo oceanico. Le conseguenze maggiori di questi cambiamenti sugli ambienti marini sono stati un incremento del livello medio dei mari ed un aumento della temperatura delle acque: questi fenomeni hanno iniziato a manifestarsi nell’Aptiano, raggiungendo il loro acme nel Cenomaniano. In un recentissimo studio, Cavin et al. (2007) hanno identificato alcuni indicatori ambientali attraverso l’analisi del record geologico e fossile con l’intento di capire
meglio i parametri che guidano l’evoluzione degli attinopterigi dal Giurassico superiore al Cretaceo superiore (intervallo di tempo in cui si realizza la diversificazione dei crown – groups Teleostei): dai loro risultati emerge che il tasso di diversificazione di molti taxa marini è direttamente proporzionale alla temperatura del mare calcolata analizzando la densità dell’O18 contenuto nello smalto dei loro denti. Questa relazione era già stata ipotizzata da Rohde (1992) in un suo studio, in cui afferma che il repentino aumento dell’energia disponibile (misurata dalla temperatura) in una popolazione di pesci marini provoca un’esplosione di diversità attraverso l’aumento del tasso di crescita della stessa popolazione ed il contemporaneo aumento del suo tasso di mutazione.
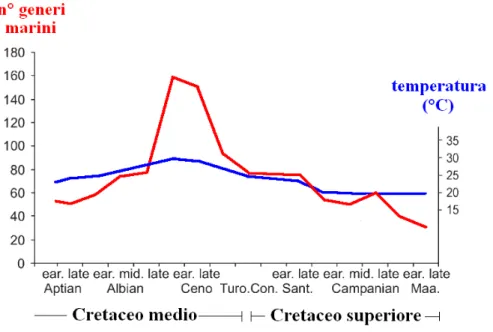
Cavin et al. (2007) sostengono che il Cretaceo medio segni l’inizio di una grande diversificazione di molti taxa teleostei marini, la quale raggiunge il suo picco massimo nel Cenomaniano (figura 3).
Figura 3. L’aumento di temperatura delle acque marine superficiali determina la diversificazione tardo cretacea dei teleostei marini (diagramma tratto da Cavin et al., 2007)
In quest’età la temperatura del mare è sempre positivamente correlata al tasso di diversificazione di alcuni gruppi giurassici come Ichthyodectiformes, Elopomorpha e Salmoniformes, ma tale correlazione diventa estremamente positiva rispetto ai nuovi gruppi come Tselfatiiformes, Aulopiformes e taxa basali degli Acanthomorpha.
Inoltre, Cavin et al. (2007) fanno un’importante considerazione: tutti i depositi a pesci cenomaniani del bacino della Tetide condividono gran parte dei taxa rivenuti; tale condivisione è meno accentuata se si paragonano tali giacimenti con le faune coeve dei giacimenti carbonatici boreali, i quali presentano molte forme endemiche. In particolare, i depositi a pesci cenomaniani del bacino
della Tetide condividono numerosi nuovi taxa, apparsi per la prima volta nel record fossile proprio durante questa età. Quindi, è molto probabile che la Tetide rappresenti un centro di origine di diversi taxa di pesci (inclusi gli acantomorfi) durante il Cretaceo, analogamente a quanto rappresenta l’area Indo – Pacifica per le forme moderne.
Il Cretaceo medio è caratterizzato anche da grandi cambiamenti ecologici che hanno determinato dei profondi stravolgimenti in molte comunità marine, come quelle bentoniche dei reefs. Questo periodo, infatti, è caratterizzato dalla grande radiazione adattativa delle rudiste, che ha consentito a questo gruppo di molluschi bivalvi di conquistare il ruolo di dominatori delle comunità di biocostruttori delle scogliere organogene a scapito delle sclerattinie, i dominatori incontrastati dei reefs terziari ed attuali.
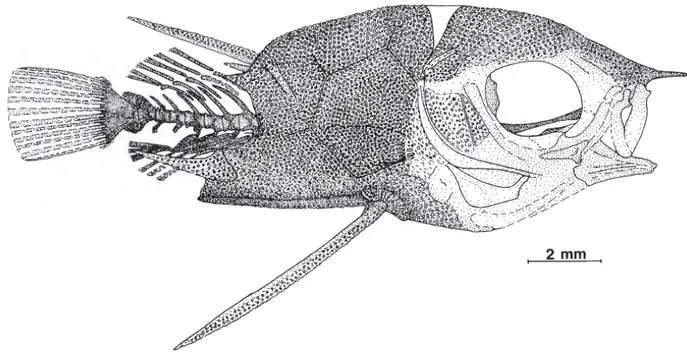
Molto probabilmente, i primi acantomorfi comparvero negli ambienti di reef tropicale, in cui la struttura del substrato è molto complessa. Le località fossilifere mesozoiche, ed in particolare quelle del bacino della Tetide, testimoniano ampiamente l’associazione tra questi pesci ed i reefs a rudiste. Nei depositi di Smoky Hill, inoltre, sono stati portati alla luce diversi reperti di acantomorfi fossilizzatisi sulla superficie interna delle grandi conchiglie di bivalvi inoceramidi; gli studiosi hanno interpretato questa associazione come l’evidenza di un rapporto mutualistico, a dimostrazione che questi pesci erano già in grado, durante il Cretaceo, di muoversi ed orientarsi in uno spazio elaborato. Osservando la struttura scheletrica di un genere acantomorfo basale del Cenomaniano come Aipichthys (figura 4), si identificano degli importanti adattamenti per incrementare la manovrabilità corporea. Le pinne pettorali di questi pesci sono posizionate sui fianchi, in modo da permettere all’animale di controllare e modulare meglio il nuoto; le pinne pelviche hanno assunto una posizione subtoracica, al di sotto delle pinne pettorali, e la loro azione impedisce il sollevamento del corpo durante le fasi di arresto; le spine dorsali ed anali, con i loro movimenti, offrono un aumento di manovrabilità, oltre che, ovviamente, un efficace sistema di difesa; il raccorciamento e la compressione laterale del corpo incidono anch’essi sull’aumento di manovrabilità. Questi elementi anatomico – funzionali costituiscono la risposta adattativa degli acantomorfi alla vita nei contesti di reef; tali caratteristiche si sono conservate nella struttura corporea degli acantomorfi per il resto del Cretaceo ed hanno certamente contribuito a sancire l’enorme successo di questo taxon a partire dalla base del Cenozoico.
Figura 4. Illustrazione di Aipichthys velifer (tratta da Patterson, 1993).
Tuttavia, Wullimann (1997) sostiene che la comparsa negli acantomorfi degli elementi anatomico – funzionali sopra citati non può prescindere dal possesso di un sistema nervoso centrale visivo capace di elaborare in modo efficace la grande mole di stimoli provenienti da un ambiente così irregolare e complesso come quello del reef. Nel suo studio sul sistema visivo pretettale mesencefalico dei teleostei, Wullimann (1997) identifica tre modelli di complessità di questa
struttura nervosa: un modello di complessità intermedia (presente nei cladi più primitivi), un secondo modello di complessità minima (presente in alcuni cladi filogeneticamente distanti tra loro e prodotto in seguito a processi indipendenti di riduzione secondaria), un terzo modello di elevata complessità (esclusivo degli acantomorfi). Se lo studio di Wullimann (1997) trovasse ulteriori conferme, il sistema visivo pretettale di complessità elevata costituirebbe una chiara sinapomorfia degli Acanthomorpha, un elemento neuro – anatomico che avrebbero dovuto possedere tutti i rappresentanti mesozoici di questo grande taxon.
Il nuovo assetto delle comunità di biocostruttori dei reefs del Cretaceo medio e superiore imposto dalla grande radiazione adattativa delle rudiste costituisce soltanto un tassello del grande mosaico di cambiamenti ecologici che hanno interessato soprattutto le comunità bentoniche marine di acque poco profonde a partire dall’ultima fase del Mesozoico fino al primo periodo dell’era cenozoica, il Paleogene (Vermeeij, 1977).
Nel Paleozoico e nella prima fase del Mesozoico, queste comunità sono sempre state dominate da organismi epifaunali o semi – infaunali. A partire dal periodo Giurassico, tuttavia, l’assetto ecologico degli ambenti marini bentonici poco profondi iniziò a mutare. Attraverso un processo di infaunalizzazione, alcuni organismi che popolavano la superficie dei fondali sabbiosi (tra cui molti echinodermi e molluschi) iniziarono a migrare dentro il substrato. Alcuni elementi epifaunali o semi – infaunali che non erano capaci di riattaccarsi al substrato una volta rimossi (come i brachiopodi ed i crinoidi) si estinsero oppure subirono una drammatica contrazione. Questi organismi furono sostituiti soprattutto dai molluschi bivalvi: alcuni di essi (ad esempio gli inoceramidi) erano in grado di aderire nuovamente al substrato attraverso la produzione del bisso; altri erano capaci di ancorarsi tenacemente al substrato attraverso la cementificazione di una delle valve (rudiste ed ostreidi). Infine, altri elementi epifaunali come i gasteropodi svilupparono progressivamente una serie di caratteristiche strutturali delle loro conchiglie per aumentarne la resistenza allo schiacciamento e contemporaneamente evolsero una serie di strutture difensive (come spine ed aperture denticolate) sulla superficie delle stesse.
Vermeeij (1977) afferma che questi grandi cambiamenti ecologici rappresentano una risposta adattativa delle prede bentoniche per opporsi al grande aumento dell’attività predatoria dei durofagi durante la seconda fase del Mesozoico ed il Paleogene. In questo intervallo, molti organismi (crostacei stomatopodi e brachiuri, molluschi gasteropodi e cefalopodi, condroitti eterodontidi e batoidei, teleostei ed uccelli) hanno evoluto la capacità di schiacciare o perforare i gusci e gli esoscheletri delle loro prede. Focalizzando l’attenzione sui Teleostei, Schaeffer & Rosen (1961) sostengono che questi pesci hanno potuto disporre di un apparato boccale che si potesse adattare a compiere svariate attività (come scalfire e schiacciare superfici dure, addentare i piccoli polipi dei
coralli, brucare le alghe) non prima del periodo Cretaceo. Se ciò che hanno affermato Schaeffer & Rosen (1961) venisse confermato da ulteriori studi ed evidenze paleontologiche, sarebbe plausibile pensare che gli acantomorfi mesozoici possedessero già un apparato boccale che consentisse loro di accedere alle molte nicchie trofiche presenti nell’ambiente bentonico.
3. 3. Radiazione cenozoica degli acantomorfi
Il record paleontologico degli acantomorfi presenta una notevole lacuna in prossimità del limite K – T. I reperti acantomorfi relativi a questo intervallo cronostratigrafico lungo circa dieci milioni di anni, che comprende l’ultimo periodo mesozoico (Maastrichtiano), il limite K – T ed il primo periodo cenozoico (Daniano), sono molto scarsi, sia numericamente che qualitativamente.
Recentemente, due località sudamericane risalenti a questo intervallo, Pajcha Pata in Bolivia centrale (Gayet et al., 2001) e Maria Farinha nel Brasile nord occidentale (Gallo et al., 2001), hanno portato alla luce resti di acantomorfi. La località di Pajcha Pata è stata datata al Maastrichtiano medio ed ha conservato i resti di una fauna marina ed estuarina; mentre il deposito fossilifero di Maria Farinha, di origine marina e molto vicino alla costa, risale al Daniano. Gli ittioliti acantomorfi di entrambe le località sono stati attribuiti all’ordine dei Perciformes; tuttavia, questi reperti si riducono a frammenti di ossa isolate, e la loro interpretazione è molto dubbia.
Il più antico Lagerstätte (termine germanico con cui si definisce un corpo sedimentario che offre un’eccezionale quantità di informazioni sulla vita del passato a causa dell’abbondanza e della qualità degli esemplari in esso contenuti) che ha portato alla luce una consistente ittiofauna del Terziario è il London Clay.
Il London Clay (“Argillite di Londra”) è una formazione geologica di origine marina risalente all’Ypresiano (Eocene inferiore, 56 – 49 milioni di anni fa); i suoi affioramenti sono localizzati nel bacino di Londra, nell’Inghilterra sud orientale. I fossili di questa formazione testimoniano la presenza di un clima subtropicale, ed i numerosi reperti vegetali (Collison, 1983) indicano che l’ambiente di sedimentazione si è sviluppato in uno spazio marino poco profondo in prossimità di una fascia costiera caratterizzata da una foresta lussureggiante. Le argilliti della formazione di London Clay sono spesse circa 150 m e si sono formate a partire da un deposito sedimentario di mare basso, profondo meno di 200 m: nella sua successione stratigrafica si contano cinque cicli
deposizionali, determinati da altrettanti eventi di trasgressione e regressione marina. Il record di teleostei di questo Lagerstätte eocenico è dominato dai generi acantomorfi: dei 60
generi di telestei rinvenuti nelle argilliti del London Clay, 47 appartengono al taxon Acanthomorpha; la percentuale relativa dei generi acantomorfi di questa ittiofauna si attesta al 78%.
Inoltre, la composizione della comunità di acantomorfi del London Clay è caratterizzata dalla
predominaza schiacciante dei generi inclusi nel crown – group Acanthomorpha,
i Percomorpha, ed in particolare di quelli che compongono l’ordine dei Perciformes. Il London Clay fornisce una delle primissime testimonianze cenozoiche della grande radiazione
evolutiva degli acantomorfi, iniziata verosimilmente nel Paleocene, protrattasi durante l’età dell’Eocene e che ha avuto come assoluto protagonista il grande sister – clade degli Smegmamorpha, in cui sono inseriti i Perciformes.
Il problema principale nell’interpretazione dei momenti chiave della storia evolutiva degli acantomorfi consiste nello stabilire quali sono gli eventi che hanno determinato la comparsa dei grandi cladi del taxon ed identificare quegli effetti provocati dalla crisi biologica al limite K – T che hanno influenzato il corso evolutivo di questo gruppo di pesci.
Fino a quando le future scoperte paleontologiche non riusciranno a colmare la lacuna nel record degli acantomorfi a cavallo del limite K – T, sarà impossibile identificare l’intervallo cronostratigrafico che sancisce l’inizio della grande diversificazione di questo taxon; questo implica che non si potrà stabilire con certezza se la grande radiazione degli acantomorfi sia stato un evento indipendente o una conseguenza della grande estinzione di massa al limite K – T.
Nel suo articolo intitolato: “Effect of the Cretaceous – Tertiary Boundary Event on Bony Fishes”, Lionel Cavin (2001) ha pubblicato i risultati dell’analisi da lui condotta sul record paleontologico di 100 famiglie di pesci ossei in corrispondenza del limite K – T: soltanto 19 famiglie (19% del totale) si sono estinte durante o alla fine del Maastrichthiano; la grande maggioranza delle famiglie estinte (80%) erano distribuite esclusivamente in ambiente marino; la quasi totalità delle famiglie estinte (90%) erano nuotatori veloci e predatori piscivori. Cavin (2001) ha ipotizzato che l’estinzione di questo tipo di pesci al limite K – T sia avvenuta in seguito ad un collasso improvviso della loro catena alimentare basata sul plancton. L’estinzione di massa del plancton in corrispondenza del limite K – T è ben documentata: ciò spiegherebbe la definitiva scomparsa di tutti quegli organismi che occupavano i livelli più alti delle catene alimentari pelagiche, fino ad arrivare ai mega predatori come alcuni grandi squali ed i mosasauri. Cavin (2001) ha affermato che questo disastro ecologico potrebbe essere stato causato da un evento improvviso, come l’impatto di un grosso meteorite: l’interruzione improvvisa della produzione primaria in seguito all’imponente sollevamento delle polveri che oscurarono l’atmosfera avrebbe prodotto una serie di eventi che disgregarono i livelli più alti della catena alimentare. In conclusione, lo studio di Cavin (2001) descrive l’estinzione dei pesci ossei al limite K – T come un evento circoscritto, poiché interessa un numero relativamente basso di famiglie, e molto selettivo, dato che colpisce le forme epipepelagiche di predatori piscivori. Il lavoro di Cavin (2001), tuttavia, presenta un punto debole: dai risultati della sua analisi non si è
rilevata alcuna estinzione di famiglie esclusivamente panctofaghe. Questo fatto può dipendere da ragioni ecologiche (non sono esistite famiglie prettamente planctofaghe in corrispondenza del limite K – T) e da ragioni tassonomiche (le eventuali famiglie esclusivamente planctofaghe non sono state prese in considerazione nell’analisi poiché le loro incerte relazioni filogenetiche mettono in dubbio il loro monofiletismo). Molti pesci planctofagi del tardo Cretaceo non sono stati presi in considerazione nello studio sopra citato proprio per quest’ultimo motivo.
Se l’ipotesi di Cavin (2001) venisse confermata, questa motiverebbe indirettamente la sopravvivenza degli acantomorfi all’estinzione cretaceo – terziaria: infatti, i membri mesozoici del taxon Acanthomorpha erano generalmente pesci di piccola taglia; la loro distribuzione si concentrava negli ambienti di mare basso; la loro struttura morfo – anatomica era adattata ad un ambiente complesso come il reef, dove era necessario possedere una buona manovrabilità corporea per nuotare in spazi stretti non solo per procacciarsi il cibo, ma anche per fuggire dalla vista dei predatori insinuandosi negli anfratti della scogliera. Per difendersi dai predatori, diversi generi di acantomorfi mesozoici, come ad esempio Protriacanthus (figura 5), avevano sviluppato un’estesa corazza di piastre dermiche; in alcuni casi, la corazza ricopriva quasi tutto il corpo.
Figura 5. Illustrazione di Protriacanthus gortanii (tratta da Tyler & Sorbini, 1996) .
Il morfotipo di acantomorfo non era certo riconducibile a quello dei veloci predatori piscivori epipelagici come gli ictiodectidi o gli encodontidi. Inoltre, la dieta degli acantomorfi del tardo Cretaceo non era stenofaga come quella dei piscivori, bensì era molto diversificata: sono state
identificate delle forme microfaghe, durofaghe, detritivore ed altre ancora. In altre parole, l’identikit di un acantomorfo del tardo Cretaceo non corrispondeva affatto a quello dei pesci che sono stati vittime dell’estinzione al limite K – T; questa grande differenza ha consentito agli acantomorfi di superare questo periodo di crisi biologica e di produrre la loro massiccia radiazione evolutiva cenozoica, ad opera quasi esclusiva dei perciformi e dei loro parenti più prossimi.
Le prime forme attribuibili con certezza all’ordine dei Perciformes compaiono in prossimità del limite K – T; i giacimenti mesozoici, tuttavia, hanno portato alla luce dei fossili che sono stati classificati proprio in quest’ordine (come quelli del genere Cylindracanthus nei giacimenti cenomaniani libanesi di Hakel e Hadjoula, oppure l’esemplare del genere Nardoichthys rinvenuto nel deposito campaniano di Nardò). Sebbene il dibattito sulla comparsa dei perciformi sia tuttora aperto, la presenza dei generi mesozoici appartenenti al taxon Percomorpha (come il Gasteroramphosus, incluso nei Gasterosteiformes, ordine incluso negli Smegmamorpha; oppure come i generi Plectocretacicus e Cretatriacanthus, collocati entrambi nei Tetraodontiformes, uno degli ordini che compone il sister – clade degli Smegmamorpha insieme ai Perciformes) potrebbero fornire delle evidenze indirette sulla presenza dei perciformi durante il Cretaceo medio e superiore. È indubbio, comunque, che l’enorme radiazione dei perciformi si sia compiuta durante il Paleocene e l’Eocene, in perfetta coincidenza con la riconquista del ruolo dominante di biocostruttori dei reefs da parte delle sclerattinie dopo l’estinzione al limite K – T delle rudiste.
I fossili del Konservat – Lagerstätte luteziano di Monte Bolca, in provincia di Verona, forniscono un’importante rappresentazione di questo punto di svolta nella storia evolutiva del taxon Acanthomorpha. La Pesciara di Bolca è uno dei siti paleontologici più famosi al mondo. Questo deposito, conosciuto da oltre cinquecento anni, consiste in un blocco di calcari litografici che si estende per poche centinaia di metri quadrati; all’interno del blocco esistono cinque livelli fossiliferi finemente stratificati. Il Lagerstätte di Monte Bolca è famoso soprattutto per i suoi pesci; tuttavia, in questo giacimento sono stati rinvenuti molti reperti di crostacei, molluschi, celenterati (estremamente rari allo stato fossile), piante acquatiche e terrestri, mentre i fossili di tetrapodi sono quasi del tutto assenti. L’assenza di vertebrati terrestri nel deposito indica che l’ambiente di sedimentazione non dovesse essere troppo vicino alla costa. I pesci di Monte Bolca rappresentano la più antica testimonianza di una moderna ittiofauna di reef tropicale. In questo sito sono presenti i membri di tutti i grandi cladi acantomorfi, comprese le famiglie più importanti dei Perciformes (Patterson, 1993); la percentuale di abbondanza relativa dei generi acantomorfi di Monte Bolca si attesta 84%, un valore molto simile a quello rilevato con i generi delle barriere coralline attuali. Le comunità di acantomorfi dei reefs cenozoici ed attuali possiedono entrambi un’elevata diversità specifica ed una relativa dominanza, in termini di specie e generi, di una o pochissime famiglie
(come quella dei Pomacentridae nei reefs attuali e la famiglia Holocentridae nei reefs eocenici come quello di Monte Bolca).
Sembra che le comunità di acantomorfi delle moderne barriere coralline si siano evolute rapidamente durante l’inizio dell’era cenozoica, restando (almeno a livello di famiglie) praticamente inalterate fino al presente. La rapida radiazione dei perciformi coincide con la riconquista del ruolo dominante nella costruzione dei reefs da parte delle sclerattinie e si realizza verosimilmente in quest’ambiente (Wullimann, 1997).
Nonostante la grande variabilità dei perciformi, tutti condividono delle caratteristiche anatomiche – funzionali molto avanzate rispetto ai teleostei più ancestrali come ad esempio i clupeiformi. Tuttavia, è necessario precisare che gran parte di queste caratteristiche osservabili nei perciformi (la posizione delle pinne pettorali e pelviche, la presenza di spine in corrispondenza delle pinne anale e dorsale, il raccorciamento e la compressione laterale del corpo) sono state ereditate dagli acantomorfi mesozoici, i quali erano già adattati alla vita nell’ambiente del reef. Un adattamento importante che si riscontra tipicamente nei perciformi è il meccanismo di protrusione della bocca attraverso il movimento in avanti del premascellare mobile. Questo sistema ha sicuramente esaltato la diversificazione dei perciformi, consentendo a questi pesci di evolvere nuove tecniche di alimentazione, di specializzare le loro diete e di accedere a nuove nicchie trofiche.
Tavola 1. Elenco delle ittiofaune a Teleostei dei depositi mesozoici esaminati in questo lavoro di tesi. Evidenziati in blu i generi che sono stati classificati nel taxon Acanthomorpha.
Hakel (Libano) Eubiodectes (†Ichthyodectiformes) Sendenhorstia (Elopiformes) Davichthys (Elopiformes) Hajoulia (Albuliformes) Lebonichthys (Albuliformes) Anguillavus (Anguilliformes) Urenchelys (Anguilliformes) Enchelion (Anguilliformes)
Ornategulum (Clupeomorpha inc. sed.) Armigatus (†Ellimmichthyiformes) Diplomystus (†Ellimmichthyiformes) Scombroclupea (Clupeiformes) Charitopsis (Gonorhynchiformes) Charitostomus (Gonorhynchiformes) Pseudoberyx (Euteleostei inc. sed.) Gaudryella (Euteleostei inc. sed.) Ginsburgia (Euteleostei inc. sed.) Exocoetoides (Aulopiformes) Telepholis (Aulopiformes) Ichthyotrynga (Aulopiformes) Apateopholis (Aulopiformes) Rhynchodercetis (Aulopiformes) Prionolepis (Aulopiformes) Enchodus (Aulopiformes) Erypholis (Aulopiformes) Phylactocephalus (Aulopiformes) Hemisaurida (Aulopiformes) Nematonotus (Aulopiformes) Sardinioides (Myctophiformes) Acrognathus (Myctophiformes) Ctenothrissa (†Ctenothrissiformes) Pharmacichthys (Pharmacichthydae) Aipichthys (Aipichthyidae)
Plesioberyx (Beryciformes inc. sed.) Caproberyx (Beryciformes)
Stichopteryx (Beryciformes)
Plectocretacicus (Tetraodontiformes)
Gasterosteiformes inc. sed.
Cylindracanthus (Acanthomorpha inc. sed.)
Hadjoula (Libano) Eubiodectes (†Ichthyodectiformes) Sendenhorstia (Elopiformes) Hajoulia (Albuliformes) Protobrama (†Plethodontidae) Anguillavus (Anguilliformes) Urenchelys (Anguilliformes) Armigatus (†Ellimmichthyiformes) Diplomystus (†Ellimmichthyiformes) Scombroclupea (Clupeiformes) Pseudoberyx (Euteleostei inc. sed.) Gaudryella (Euteleostei inc. sed.) Ginsburgia (Euteleostei inc. sed.) Exocoetoides (Aulopiformes) Telepholis (Aulopiformes) Ichthyotrynga (Aulopiformes) Rhynchodercetys (Aulopiformes) Prionolepis (Aulopiformes) Enchodus (Aulopiformes) Erypholis (Aulopiformes) Phylactocephalus (Aulopiformes) Nematonotus (Aulopiformes) Sardinioides (Myctophiformes) Acrognathus (Myctophiformes) Ctenothrissa (†Ctenothrissiformes) Heterothrissa (†Ctenothrissiformes) Pateroperca (†Pattersonichthyidae) Pattersonichthys (†Pattersonichthydae) Phoenicolepis (†Pattersonichthydae) Humilichthys (†Pattersonichthydae) Freigichthys (†Aipichthyioidea) Aipichthys (†Aipichthyidae) Pycnosteroides (†Pycnosteroididae) Cryptoberyx (Beryciformes inc. sed.) Stichocentrus (Beryciformes) Stichopteryx (Beryciformes) Lissoberyx (Beryciformes) Microcapros (Beryciformes) Libanoberyx (Beryciformes) Hgulichthys (Beryciformes)