Contents lists available atScienceDirect
Phytomedicine
journal homepage:www.elsevier.com/locate/phymed
Original Article
The major Boswellia serrata active 3-acetyl-11-keto-
β-boswellic acid
strengthens interleukin-1
α upregulation of matrix metalloproteinase-9 via
JNK MAP kinase activation
Elia Ranzato
a, Simona Martinotti
a, Andrea Volante
b, Aldo Tava
c, Maria Angela Masini
a,
Bruno Burlando
d,e,*
aDipartimento di Scienze e Innovazione Tecnologica (DISIT), Università del Piemonte Orientale, viale T. Michel 11, 15121, Alessandria, Italy
bConsiglio per la Ricerca in Agricoltura e l'Analisi dell'Economia Agraria - Rice Research Unit (CREA-RIS), SS 11 per Torino, km 2.5, 13100, Vercelli, Italy cConsiglio per la Ricerca in Agricoltura e l'Analisi dell'Economia Agraria - Centro di Ricerca Zootecnia e Acquacoltura (CREA-ZA), Viale Piacenza 29, 26900, Lodi, Italy dDipartimento di Farmacia, Università di Genova, Viale Benedetto XV 3, 16132, Genova, Italy
eIstituto di Biofisica, CNR, via De Marini 6, 16149, Genova, Italy
A R T I C L E I N F O
Keywords: Extracellular matrix Gelatin zymography HaCat keratinocytes MAP kinases NF-κB, skin inflammationA B S T R A C T
Background: Boswellia serrata gum resin has attracted pharmacological interest as an alternative anti-nflammatory.
Purpose: We studied the application of an ethanolic extract of the resin and its main active 3-O-acetyl-11-keto-β-boswellic acid (AKBA) against inflammatory degeneration of skin extracellular matrix.
Study design: We compared the effects of the extract and AKBA on the activity of MMP-2 and MMP-9 (72-kDa and 92-kDa type IV collagenases) in HaCaT keratinocytes exposed to interleukin-1α (IL-1α) as a skin in-flammation model.
Methods: MMP activity in cell conditioned medium was assayed by gelatin zymography, while NF-KB and MAP
kinase activations were evaluated by Western blotting.
Results: IL-1α (10 ng/ml) upregulated MMP-9 but not MMP-2 in HaCaT cells. The extract, used at 2.3, 4.6 and 9.3 µg/ml, had no effect, but in combination with IL-1α showed MMP-9 inhibition at the lowest dose and in-creased upregulation at the highest one. AKBA alone, at the same concentrations (corresponding to 5, 10, and 20 µM), did not stimulate MMP-9, but together with IL-1α induced an increased upregulation at the lowest dose that progressively disappeared at higher doses. WB analysis showed that IL-1α induced phosphorylation of NF-κB p65, while AKBA abolished this effect at 20 µM, but conversely increased it at 5 µM. Screening of MAP kinase phosphorylation showed a combined activation of IL-1α/AKBA on JNK, while the JNK inhibitor SP600125 abolished MMP-9 upregulation induced by IL-1α/AKBA.
Conclusion: The enhancing effect of IL-1α/AKBA on MMP-9 at low AKBA concentration seems to involve the activation of JNK-mediated NF-κB pathway. Conversely, the extract inhibits the IL-1α effect at low doses, but not at higher ones, where AKBA and possibly otherβ-boswellic acids reach concentrations that potentiate the effect of IL-1α. The extract at low doses could protect the skin against degenerative processes of extracellular matrix, while keto-β-boswellic acids seem unsuitable for this purpose.
Introduction
The gum resin from the tropical bush Boswellia serrata Roxb. ex Colebr. (Burseraceae), known as Indian frankincense, is a complex mixture of terpenes and polysaccharides used as traditional Ayurvedic remedy for chronic inflammatory diseases. The resin has attracted
pharmacological interest for therapeutic potentials against arthritis, asthma, colitis, Crohn's disease, and cancer, and is considered a pro-mising alternative to nonsteroidal antinflammatory drug (Siddiqui, 2011). Its main active principles are pentacyclic triterpenes known as boswellic acids, particularly 3-O-acetyl-11-keto-β-boswellic acid (AKBA), which has been shown to exert antinflammatory activity
http://dx.doi.org/10.1016/j.phymed.2017.09.010
Received 14 January 2017; Received in revised form 13 June 2017; Accepted 24 September 2017
*Corresponding author.
E-mail address:[email protected](B. Burlando).
Abbreviations: AKBA, 3-O-acetyl-11-keto-β-boswellic acid; ERK, extracellular signal–regulated kinase; IL, interleukin; JNK, c-Jun N-terminal kinase; KBA, 11-keto-β-boswellic acid; MMP, matrix metalloproteinase; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells
0944-7113/ © 2017 Elsevier GmbH. All rights reserved.
(Altmann et al., 2002; Sailer et al., 1996), and pro-apoptotic effects on cancer cells (Liu et al., 2002; Xia et al., 2005).
Indian frankincense has been also used along history in skin care treatments (Diamandopoulos, 1996; Lardos, 2006), while there is a growing interest for its possible skin antinflammatory and antiage ef-fects (Gi and Heon, 2004; Seong and Joon, 2006). An important process of skin inflammation and aging is extracellular matrix remodeling, where matrix metalloproteinases (MMPs) are known to play a pivotal role. These latter are a family of zinc-dependent endopeptidases pro-duced by different tissues, including skin cells, which are capable of degrading various extracellular matrix components ( Birkedal-Hansen et al., 1993). MMPs are induced in response to various cyto-kines, growth factors, cell-matrix interactions, and altered cell-cell contacts, while evidence is accumulating about their involvement in morphogenesis, angiogenesis, and tissue repair. However, MMPs also play pathogenetic roles in osteoarthritis, rheumatoid arthritis, chronic ulcers, photoaging, and periodontitis, as well as in tumor cell invasion and metastasis (Kahari and Saarialho-Kere, 1997).
Inhibitory effects of Indian frankincense and its constituents on MMPs have been reported. It has been shown that AKBA inhibits MMP-3 activity in endotelial tissue, while a mixture of acetyl-α-boswellic and acetyl-β-boswellic acids has inhibited the secretion of MMP-2 and MMP-9 in human fibrosarcoma cells (Roy et al., 2006; Zhao et al., 2003). However, scarce information is available about possible effects of these compounds on skin cells and the underlying molecular me-chanisms.
The problem of different and even opposite effects between phyto-complex and its single constituents is well known (Wagner, 2005). The cytotoxicity of B. serrata extract and its constituent AKBA on skin cells, including the spontaneously immortalized keratinocyte cell line HaCaT, has been previously studied (Burlando et al., 2008). The present study is aimed at elucidating the effect of the same agents on the activity of MMP-2 and MMP-9 (72-kDa and 92-kDa type IV collagenases, respec-tively) in HaCaT cells exposed to interleukin-1α (IL-1α) as a model of skin inflammation. IL-1α is involved in inflammation, differentiation and other processes of keratinocytes, and has been reported to stimulate the secretion and activity of MMPs, particularly MMP-9 (Han et al., 2005). In order to explore the regulatory pathways involved in these effects, we focused on the proinflammatory factor NF-ĸB and on mi-togen-activated protein kinases (MAPKs), which are both known to be involved in the regulation of MMPs in keratinocytes (Cooper et al., 2004; McCawley et al., 1999).
Materials and methods Plasticware and reagents
Cell culture flasks and plates were obtained from Costar (Cambridge, MA, USA). Standard culture medium, foetal bovine serum (FBS) and supplemented compounds were purchased from Sigma–Aldrich (St Louis, MO, USA). TripLE Express, Penicillin, Streptomycin, and Glutamine were purchased from Gibco (Invitrogen Life Technologies, Milan, Italy). An ethanolic extract of B. serrata gum resin was purchased from EPO, Istituto Farmochimico Fitoterapico (cat. n. 3536765, Milan, Italy), while 3-O-acetyl-11-keto-β-boswellic acid (AKBA) was from Alexis Corp. (Lausen, Switzerland). The inhibitors SB203580, PD98059, and SP600125 were from Calbiochem (La Jolla, CA, USA). All other chemicals of analytical grade were from Sigma. Qualitative and quantitative evaluation of boswellic acids
Quantitation of bioactive compounds in the B. serrata gum resin extract was carried out using a gas chromatography-flame ionization detector (GC–FID) of the corresponding methyl-silyl derivatives using hederagenin as internal reference. Gas chromatography-mass spectro-metry (GC–MS) was also used for identification of the compounds,
comparing results with those from authentic reference standards and data available from literature (Hairfield et al., 1989; Mathe et al., 2004). Some 5 mg of sample, to which 0.5 mg of hederagenin was added as internal standard, were suspended in 2 ml of methanol, so-nicated for 1 min and treated with CH2N2 for 10 min at room
tem-perature. After solvent evaporation under stream of nitrogen, the solid was treated with 0.5 ml of pyridin/hexamethyldisilazane/chloro-trimethylsilane 2:1:1 and heated at 70 °C for 10 min. After cooling, 0.5 ml of isooctane was added and samples used (1 µl) for GC analyses. Stock solution of hederagenin (5.2 mg/ml), β-boswellic acid (2.5 mg/ml) and AKBA (2.0 mg/ml) were prepared in butanol. Calibration curves were determined using a series of standard solutions containing a constant amount of internal standard (hederagenin) with varying amounts of pure compounds. All the prepared standard solu-tions were methylated-silylated as reported and then injected in GC in quadruplicate for each concentration. A linear response (conc./int. std conc.) vs. peak area (area/int. std area) was obtained between 0.011–0.468 mg/ml and 0.012–0.472 mg/ml for β-boswellic acid and AKBA, respectively. The limit of detection was evaluated as 0.005 and 0.006 mg/ml forβ-boswellic acid and AKBA, respectively. Intra-day and inter-day variations were recorded to determine the precision of the developed analytical method. The obtained regression equations and relative correlation coefficients were: y = 0.9448x−0.0354, R2= 0.9950 and y = 1.2143x−0.0043, R2= 0.9943 for β-boswellic acid and AKBA, respectively. Quantitations ofα- and β-boswellic acids, and their 3-O-acetyl- derivatives were referred to theβ-boswellic acid calibration curve, while quantitations of 11-keto-β-boswellic acid and AKBA were referred to the AKBA calibration curve.
GC–FID analyses were carried out using a Perkin–Elmer (Milano, Italy) model 8500 GC equipped with a 30 m × 0.32 mm i.d., 0.25 µm, DB-5 capillary column. Injector and detector temperatures were set at 320 and 330 °C, respectively. The oven temperature program was as follows: 50 °C for 2 min, increased at 15 °C/min to 230 °C and then in-creased at 4 °C/min to 330 °C for 10 min. Samples (0.5 µl) were injected in the‘splitless’ mode. The carrier gas was He with a head pressure of 18.0 psi (2.0 ml/min).
GC–MS analyses were carried out using a Perkin-Elmer Clarus 500 GC equipped with a MS detector and a 30 m × 0.25 mm i.d., 0.25 µm, Elite-5MS capillary column using the same chromatographic conditions as for GC–FID. Mass spectra were acquired over the 50–800 amu range at 1 scan/s with an ionizing electron energy of 70 eV. Transfer line temperature was 300 °C, and the carrier gas was He at 1.2 ml/min. Cell cultures
The spontaneously immortalized HaCaT keratinocytes (Boukamp et al., 1998) were maintained and cultured in DMEM medium supple-mented with 10% FBS, 100 U/ml penicillin, 100 mg/ml streptomycin, and 290 mg/ml glutamine, at 37 °C in humidified atmosphere con-taining 5% CO2. All cell cultures proved mycoplasma-free on periodic
checks using the DNAfluorochrome staining method with bis-benzy-mide dye (Hoechst 33258).
Cell treatments for gelatin zymography
Cells were seeded 24 h before treatment in 6-well plates at 3.5 × 105 cells/well. The tested compounds (B. serrata extract and AKBA) were preliminary dissolved in DMSO and diluted in DMEM medium supplemented as above (vehiclefinal concentration < 0.5%). Treatments were performed for 48 h using 2.3 µg/ml, 4.6 µg/ml, or 9.3 µg/ml B. serrata extract or AKBA, with or without 10 ng/ml IL-1α (Sigma). Control cultures received the same concentration of vehicle. For pharmacological inhibition assays, we treated the cells for 48 h with PD98059 (ERK inhibitor, 10 µM), SB203580 (p38 inhibitor, 20 µM), and SP600125 (JNK Inhibitor, 100 µM) in DMEM medium.
The conditioned medium was then collected and concentrated
fold using Centricon filters (Millipore, Bedford, MA, USA) with a 30 KDa cut-off, by centrifugation for 30 min at 2000 rpm. Samples were then stored at–20 °C until use. The corresponding cells were lysed with NP-40 buffer (50 mM Tris/HCl, pH 8.0, 150 mM NaCl, 5 mM EDTA, Protease Inhibitors Cocktail by Sigma–Aldrich, 1% (v/v) NP-40); the lysate was thoroughly homogenised by passing through a syringe needle. Total protein concentration was determined using the bicinch-oninic acid-based BCA Protein Assay Kit (Rockford, IL, USA), following the manufacturer's protocol.
Gelatin zymography
Concentrated media processed as described above were mixed with an equal volume of a 2 × loading buffer (0.4 M Tris–HCl pH 6.8, 20% glycerol, 5% SDS, a pinch of bromophenol blue) and then loaded onto a non-denaturing 6% polyacrylamide gel containing 0.1% (w/v) gelatin. In order to use volumes corresponding to comparable amounts of cells, the volumes of each sample were adjusted according to the protein concentrations of the corresponding cell lysate. Proteins were resolved by electrophoretic run at 150 V for 90 min. After electrophoresis, the SDS in the gel was removed by incubation with 2.5% Triton X-100 for 1 h. The gelatinolytic activity was developed in a buffer containing Fig. 1. Chemical structure of boswellic acids detected by GC–MS in the gum resin extract of B. serrata: α-boswellic acid (αBA); β-boswellic acid (βBA); acetyl-α-boswellic acid (AαBA); 3-acetyl-β-boswellic acid (AβBA); 11-keto-β-boswellic acid (KBA); 3-acetyl-11-keto-β-boswellic acid (AKBA).
5 mM CaCl2, 200 mM NaCl, and 50 mM Tris, pH 7.5, by incubation for
18 h at 37 °C. The enzymatic activity was then visualized by staining in 0.5% Coomassie Brilliant Blue G-250, 10% glacial acetic acid, 30% methanol. Gel images were acquired with a GS-710 densitometer and the Quantity One software (Bio Rad, Hercules, CA, USA), and analysed using the ScionImage 4.0.3.2 software (Scion Corp., USA). Experiments were carried out in triplicate.
Western blot
Cells were seeded in 12-well plates, exposed for 24 h to various treatments as indicated, and then lysed with 200 µl/well of NP-40 buffer, supplemented with 5 mM NaF, 1 mM sodium orthovanadate, 3 mM sodium pyrophosphate and Protease Inhibitors Cocktail by Sigma–Aldrich. The lysate was thoroughly homogenised by passing through a syringe needle. Total protein concentration was determined as above. Samples were denaturated by adding an equal volume of 2 × Laemmli Buffer (0.25 mM Tris–HCl pH 6.8, 0.4% SDS, 2% glycerol, 2%β-mercaptoethanol, a pinch of bromophenol blue) and boiling for 5–10 min. Volumes corresponding to 50 µg of total proteins were then resolved by electrophoresis on a 10% polyacrylamide gel. A molecular size marker (Precision Plus Protein Std All Blue, Bio-Rad, Hercules, CA, USA) was included as a reference. Proteins were then transferred onto a nitrocellulose membrane using a Bio-Rad Mini Trans Blot electro-phoretic transfer unit. The membranes were blocked for nonspecific protein with 5% nonfat, dry milk in PBS and then probed with specific rabbit anti-human primary antibodies (1:1000) against ERK1/2, p-p38, p-p65, p-JNK, and the corresponding non-phosphorylated forms (Cell Signaling Technology, Celbio SpA, Milan, Italy), at room tem-perature for 1 h, or at 4 °C overnight. Membranes were then washed three times (10 min per wash) with PBS supplemented with 0.05% Tween-20, to remove unbound antibodies, and then further incubated with appropriate horseradish-peroxidase-conjugated secondary anti-bodies. Membranes were developed by an ECL kit (Millipore, Billerica, MA, USA), according to the manufacturer's protocol, digitized with the Quantity One Image Software (ChemiDoc XRS, Bio-Rad laboratories, Hercules, CA) and normalized against proper loading controls. Band intensities were quantified using the ScionImage 4.0.3.2 software (Scion Corp., USA).
Statistics
Data were analysed using the Instat and Prism software package (GraphPad Software, Inc).
Results and discussion
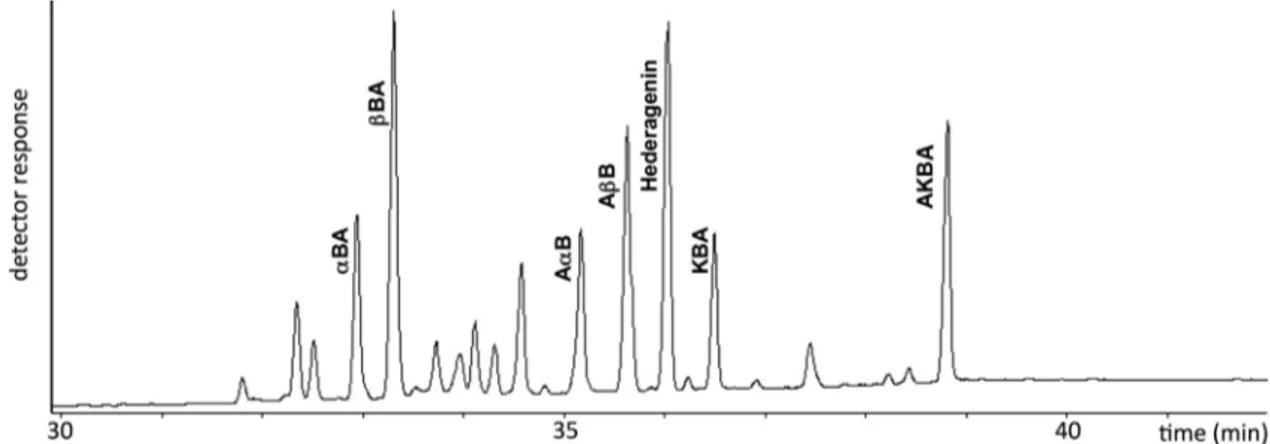
The chemical structures of the identified boswellic acids are re-ported inFig. 1, while their gas chromatographic separation is shown in
Fig. 2. All compounds in the complex mixture were baseline separed. Analyses were carried out using hederagenin as internal standard. This compound has a suitable retention time without interferences with other peaks. Quantitative results (mg/g dry matter) of the identified boswellic acids are reported inTable 1, as means of three independent determinations ± standard deviation.
By using gelatin zymography analysis (Leber and Balkwill, 1997), we evaluated the activity of two major gelatinases, namely MMP-2 and MMP-9, in the conditioned medium of HaCaT keratinocytes after 48 h incubations. The bands corresponding to these gelatinases in their ac-tive and inacac-tive form had been previously identified (Ranzato et al., 2011), while in the present study, their identity was confirmed by comparison with standard homologs obtained from HL-60 cells stimu-lated with phorbol 12-myristate 13-acetate (seeFig. 5).
Inflammatory activation of HaCaT cells, simulated by exposure to 10 ng/ml IL-1α, induced an increase of MMP-9 activity, especially the pro-enzyme form (which is active in zymography) (Fig. 3A). In contrast, MMP-2 showed no induction by IL-1α, consistent with what observed bySalo et al. (1994). In experiments with B. serrata extract and AKBA, we used subtoxic concentrations, i.e. lower than previously-determined IC05values (Burlando et al., 2008). The extract, used alone at different
concentrations (2.3, 4.6 and 9.3 µg/ml), was not able to induce any increase in MMP-9 activity. However, when used in combination with IL-1α, it was able to inhibit MMP-9 stimulation at the lowest con-centration, whereas, at higher concentrations the induction of MMP-9 progressively increased, up to a level significantly higher than that in-duced by IL-1α alone, thus revealing a combined effect between the extract and the cytokine (Fig. 3A).
The use of AKBA at the same concentrations of the extract (corre-sponding to 5, 10, and 20 µM AKBA) did not stimulate MMP-9, but in contrast there was an inhibitory effect at the highest concentration. However, a strong combined effect with IL-1α, suggesting a synergistic interaction, was observed. Unlike the extract, the combined IL-1α/ AKBA effect occurred at the lowest AKBA concentration (Fig. 3B). At higher AKBA concentrations the combined effect on MMP-9 progres-sively decreased, but MMP-9 induction did not fall below that of IL-1α
Fig. 2. Gas chromatogram of B. serrata gum resin extract (abbreviations as inFig. 1). Table 1
Quantitative evaluation of boswellic acids in the B. serrata gum resin extract expressed as mg/g dry matter.
Compound mg/g α-boswellic acid 37.5 ± 0.52 β-boswellic acid 81.2 ± 2.02 3-O-acetyl-α-boswellic acid 36.5 ± 0.27 3-O-acetyl-β-boswellic acid 62.8 ± 1.34 11-keto-β-boswellic acid 37.5 ± 1.25 AKBA 64.3 ± 1.51 Total 319.8 ± 6.43
alone (Fig. 3B).
With the aim of disclosing the mehanism of the biphasic MMP-9 induction due to combination of IL-1α and AKBA, we made a check for the activation of such regulatory factors as NF-ĸB and MAP kinases, which are known to act upstream of MMP-9 induction in inflammatory processes (Wuertz et al., 2012). Cell responses were recorded after 24 h incubations with 10 ng/ml IL-1α, 5 µM AKBA, or a combination thereof. NF-ĸB plays a fundamental role in the development and progression of inflammatory diseases such as arthritis, asthma, atherosclerosis, and gut inflammation (Baldwin, 2001; Tak and Firestein, 2001). A mole-cular marker of its activation is p65 subunit phosphorylation, revealing translocation to the nucleus (Buss et al., 2004). NF-ĸB is considered to be a target of AKBA, which has been shown to inhibit NF-ĸB signalling in different proinflammatory and stress conditions (Ammon, 2016; Catanzaro et al., 2015; Syrovets et al., 2005).
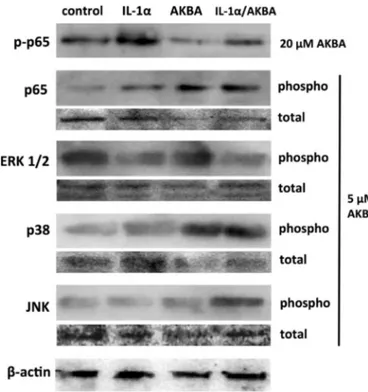
In our experiments, as expected, IL-1α induced p65
phosphorylation, while AKBA at a concentration of 20 µM reduced basal p65 phosphorylation level and abolished the effect of IL-1α (Fig. 4). This is consistent with literature data showing that IL-1α is an activator of NF-ĸB (Xu et al., 1997), and that this nuclear factor induces the transcription of the MMP-9 gene (Eberhardt et al., 2000). However, quite unexpectedly, a dose of 5 µM AKBA increased p65 phosphoryla-tion, while the combination of IL-1α and AKBA also produced an in-crease, although not exceeding that of AKBA alone (Fig. 4).
Analysis of the phosphorylation status of main MAP kinases showed no effect of AKBA on ERK1/2, while IL-1α reduced its phosphorylation below control levels (Fig. 4). IL-1α was uneffective on p38, while AKBA induced its phosphorylation, but no interaction between the two agents was observed (Fig. 4). Conversely, a combined stimulatory effect of IL-1α/AKBA was visible on JNK, as neither IL-1α nor AKBA induced phosphorylation when used alone, but a significant induction was ob-served with their combination (Fig. 4).
We made an attempt at correlating Western blot data with zimo-graphy by using the MAP kinase inhibitors PD98059 (ERK1/2 inhibitor, 10 µM), SB203580 (p38 inhibitor, 20 µM), and SP600125 (JNK in-hibitor, 10 µM). Zymograms were obtained from cells exposed for 48 h to 10 ng/ml IL-1α together with 5 µM AKBA, in the absence or presence of each inhibitor. By this way, we observed the above-described, strong increase of MMP-9 activity after exposure to IL-1α/AKBA combination. All inhibitors reduced this stimulatory effect, but the strongest reduc-tion occurred with the JNK inhibitor (Fig. 5).
Various studies have established a link between JNK activation and MMP-9 induction (Cheng et al., 2016; Hsieh et al., 2014), while it has also been shown that the activation of NF-κB is mediated by JNK phosphorylation as an upstream signal (Borsello and Forloni, 2007). Based on these literature reports, the complex of our data indicates that the synergistic effect of IL-1α and AKBA at low concentration involves the activation of the JNK-mediated, NF-κB-dependent pathway, thereby increasing MMP-9 expression. At higher AKBA concentrations, an in-hibitory effect on NF-κB would become prevalent, thus explaining the disappearance of the synergism.
Fig. 3. Gelatinolytic activity of MMP-9 in HaCaT cells, after 48 h treatment with B. serrata extract (panel A) or AKBA (panel B), with and without stimulation with 10 ng/ml IL-1α (grey and white bars, respectively). Data represent band intensity mean values ± sd ob-tained by image analysis of 3 independent experiments (one of which is shown at the bottom). Different letters on bars indicate significant differences among groups (p < .01) according to the Tukey's test.
Fig. 4. Western blot analyses showing the effects of 10 ng/ml IL-1α, AKBA, or their combination on the activation state of NFKB-p65, ERK1/2, p38 and JNK. Total cell lysates (40 µg protein⁄lane) were separated on 12% SDS–-PAGE, transferred onto a nitrocellulose membrane, labelled with antibodies against phospho NF-KB p65, ERK1/2, p38, and JNK, and then stripped and reprobed with anti-actin as loading control.
As for the opposite dose-response effect exerted on MMP-9 by the IL-1α/extract combination with respect to IL-1α/AKBA, the explanation could reside in the amounts of AKBA and its second major allied 11-keto-β-boswellic acid (KBA) in the extract. GC analysis revealed average AKBA and KBA contents of 64.3 and 37.5 mg/g, respectively (i.e. taken together about 10%) (Table 1). Hence, when used at 2.3 µg/ ml, the extract contains 0.23 µg/ml keto-β-boswellic acids, and its in-hibition on MMP-9 stimulation by IL-1α should be attributed to other constituents. Conversely, in treatments with IL-1α and 9.3 µg/ml ex-tract, showing enhanced induction of MMP-9 activity, keto-β-boswellic acids reach a concentration of 0.95 µg/ml, i.e. close to the treatment with IL-1α and 2.3 µg/ml AKBA (5 µM), where a combined stimulatory effect is observed.
Conclusions
In conclusion, this study provided useful indications for Indian frankincense application to skin problems. The extract could effectively inhibit MMP-9 stimulation, acting as a protective agent for skin extra-cellular matrix against inflammatory and degenerative processes. However, in order to achieve such an alleviating effect proper dosage should be used in formulations, by keeping keto-β-boswellic acids below levels of 0.5 µg/ml. On the contrary, purified keto-β-boswellic acids seem to be unsuitable as a remedy against degenerative processes of the skin extracellular matrix.
Conflict of interest
The authors report no conflict of interest. Acknowledgments
This study was funded by a grant from Università del Piemonte Orientale “Ricerca Locale 2011-BB”. ER was recipient of a Research Fellowship from the University of Piemonte Orientale. SM was recipient of a PhD scholarship from the Italian Government.
References
Altmann, A., Fischer, L., Schubert-Zsilavecz, M., Steinhilber, D., Werz, O., 2002. Boswellic acids activate p42(MAPK) and p38 MAPK and stimulate Ca(2+) mobilization. Biochem. Biophys. Res. Commun. 290, 185–190.
Ammon, H.P., 2016. Boswellic acids and their role in chronic inflammatory diseases. Adv. Exp Med. Biol. 928, 291–327.
Baldwin, A.S., 2001. Series introduction: the transcription factor NF-kappaB and human disease. J. Clin. Invest. 107, 3–6.
Birkedal-Hansen, H., Moore, W.G., Bodden, M.K., Windsor, L.J., Birkedal-Hansen, B., DeCarlo, A., Engler, J.A., 1993. Matrix metalloproteinases: a review. Crit. Rev. Oral. Biol. Med. 4, 197–250.
Borsello, T., Forloni, G., 2007. JNK signalling: a possible target to prevent neurodegen-eration. Curr. Pharm. Des. 13, 1875–1886.
Boukamp, P., Petrussevska, R.T., Breitkreutz, D., Hornung, J., Markham, A., Fusenig, N.E., 1998. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 106, 761–771.
Burlando, B., Parodi, A., Volante, A., Bassi, A.M., 2008. Comparison of the irritation potentials of Boswellia serrata gum resin and of acetyl-11-keto-beta-boswellic acid by in vitro cytotoxicity tests on human skin-derived cell lines. Toxicol. Lett. 177, 144–149.
Buss, H., Dorrie, A., Schmitz, M.L., Hoffmann, E., Resch, K., Kracht, M., 2004. Constitutive and interleukin-1-inducible phosphorylation of p65 NF-{kappa}B at serine 536 is mediated by multiple protein kinases including I{kappa}B kinase (IKK)-{alpha}, IKK{beta}, IKK{epsilon}, TRAF family member-associated (TANK)-binding kinase 1 (TBK1), and an unknown kinase and couples p65 to TATA-binding protein-associated factor II31-mediated interleukin-8 transcription. J. Biol. Chem. 279, 55633–55643.
Catanzaro, D., Rancan, S., Orso, G., Dall'Acqua, S., Brun, P., Giron, M.C., Carrara, M., Castagliuolo, I., Ragazzi, E., Caparrotta, L., Montopoli, M., 2015. Boswellia serrata preserves intestinal epithelial barrier from oxidative and inflammatory damage. PLoS One 10, e0125375.
Cheng, H.L., Hsieh, M.J., Yang, J.S., Lin, C.W., Lue, K.H., Lu, K.H., Yang, S.F., 2016. Nobiletin inhibits human osteosarcoma cells metastasis by blocking ERK and JNK-mediated MMPs expression. Oncotarget 7, 35208–35223.
Cooper, K.L., Myers, T.A., Rosenberg, M., Chavez, M., Hudson, L.G., 2004. Roles of mi-togen activated protein kinases and EGF receptor in arsenite-stimulated matrix me-talloproteinase-9 production. Toxicol. Appl. Pharmacol. 200, 177–185.
Diamandopoulos, A.A., 1996. Organic and inorganic cosmetics in the preclassical Eastern Mediterranean. Int. J. Dermatol. 35, 751–756.
Eberhardt, W., Huwiler, A., Beck, K.F., Walpen, S., Pfeilschifter, J., 2000. Amplification of IL-1 beta-induced matrix metalloproteinase-9 expression by superoxide in rat glo-merular mesangial cells is mediated by increased activities of NF-kappa B and acti-vating protein-1 and involves activation of the mitogen-activated protein kinase pathways. J. Immunol. 165, 5788–5797.
Gi, C.B., Heon, J.J., 2004. Cosmetic composition for enhancing skin elasticity comprising Boswellia extract as active ingredient to remove skin wrinkles and enhance skin elasticity. Pat n KR20040078498.
Hairfield, E.M., Hairfield, H.H., McNair, H.M., 1989. GC, GC/MS, and TLC of β-boswellic acid and O-acetyl-β-boswellic acid from B. serrata, B. carteii, and B. papyrifera. J. Chromatogr. Sci. 27, 127–133.
Han, Y.P., Downey, S., Garner, W.L., 2005. Interleukin-1alpha-induced proteolytic acti-vation of metalloproteinase-9 by human skin. Surgery 138, 932–939.
Hsieh, S.C., Tsai, J.P., Yang, S.F., Tang, M.J., Hsieh, Y.H., 2014. Metformin inhibits the invasion of human hepatocellular carcinoma cells and enhances the chemosensitivity to sorafenib through a downregulation of the ERK/JNK-mediated NF-kappaB-de-pendent pathway that reduces uPA and MMP-9 expression. Amino Acids 46, 2809–2822.
Kahari, V.M., Saarialho-Kere, U., 1997. Matrix metalloproteinases in skin. Exp. Dermatol. 6, 199–213.
Lardos, A., 2006. The botanical materia medica of the Iatrosophikon–a collection of prescriptions from a monastery in Cyprus. J. Ethnopharmacol. 104, 387–406.
Leber, T.M., Balkwill, F.R., 1997. Zymography: a single-step staining method for quan-titation of proteolytic activity on substrate gels. Anal. Biochem. 249, 24–28.
Liu, J.J., Nilsson, A., Oredsson, S., Badmaev, V., Duan, R.D., 2002. Keto- and acetyl-keto-boswellic acids inhibit proliferation and induce apoptosis in Hep G2 cells via a cas-pase-8 dependent pathway. Int. J. Mol. Med. 10, 501–505.
Mathe, C., Culioli, G., Archier, P., Vieillescazes, C., 2004. Characterization of archae-ological frankincense by gas chromatography-mass spectrometry. J. Chromatogr. A 1023, 277–285.
McCawley, L.J., Li, S., Wattenberg, E.V., Hudson, L.G., 1999. Sustained activation of the mitogen-activated protein kinase pathway. A mechanism underlying receptor tyr-osine kinase specificity for matrix metalloproteinase-9 induction and cell migration. J. Biol. Chem. 274, 4347–4353.
Ranzato, E., Martinotti, S., Volante, A., Mazzucco, L., Burlando, B., 2011. Platelet lysate modulates MMP-2 and MMP-9 expression, matrix deposition and cell-to-matrix ad-hesion in keratinocytes andfibroblasts. Exp. Dermatol. 20, 308–313.
Roy, S., Khanna, S., Krishnaraju, A.V., Subbaraju, G.V., Yasmin, T., Bagchi, D., Sen, C.K., 2006. Regulation of vascular responses to inflammation: inducible matrix metallo-proteinase-3 expression in human microvascular endothelial cells is sensitive to an-tiinflammatory Boswellia. Antioxid. Redox Signal 8, 653–660.
Sailer, E.R., Subramanian, L.R., Rall, B., Hoernlein, R.F., Ammon, H.P., Safayhi, H., 1996. Acetyl-11-keto-beta-boswellic acid (AKBA): structure requirements for binding and 5-lipoxygenase inhibitory activity. Br. J. Pharmacol. 117, 615–618.
Salo, T., Makela, M., Kylmaniemi, M., Autio-Harmainen, H., Larjava, H., 1994. Expression of matrix metalloproteinase-2 and -9 during early human wound healing. Lab Invest 70, 176–182.
Seong, N.H., Joon, K.Y., 2006. Cosmetic compositions for skin care containing extract of Boswellia carterii Birdw. Pat n KR20060072532.
Siddiqui, M.Z., 2011. Boswellia serrata, a potential antiinflammatory agent: an overview. Indian J. Pharm. Sci. 73, 255–261.
Syrovets, T., Gschwend, J.E., Buchele, B., Laumonnier, Y., Zugmaier, W., Genze, F., Simmet, T., 2005. Inhibition of IkappaB kinase activity by acetyl-boswellic acids promotes apoptosis in androgen-independent PC-3 prostate cancer cells in vitro and in vivo. J. Biol. Chem. 280, 6170–6180.
Tak, P.P., Firestein, G.S., 2001. NF-kappaB: a key role in inflammatory diseases. J. Clin. Invest. 107, 7–11.
Wagner, H., 2005. Natural products chemistry and phytomedicine in the 21st century:
Fig. 5. Gelatinolytic activities of the conditioned medium of HaCaT cells incubated or not with a combination of IL-1α and AKBA, in the presence of each of the following: none (–), ERK1/2 inhibitor PD98059 (PD), p38 inhibitor SB203580 (SB), and JNK inhibitor SP600125 (SP). Standard MMP-9 and MMP-2 (MW) were obtained from serum-free medium conditioned by HL60 cells stimulated for 24 h with 100 nM phorbol 12-myristate 13-acetate.
new developments and challenges. Pure Appl. Chem. 77, 1–6.
Wuertz, K., Vo, N., Kletsas, D., Boos, N., 2012. Inflammatory and catabolic signalling in intervertebral discs: the roles of NF-kappaB and MAP kinases. Eur. Cell Mater. 23, 103–119 discussion 119-120.
Xia, L., Chen, D., Han, R., Fang, Q., Waxman, S., Jing, Y., 2005. Boswellic acid acetate induces apoptosis through caspase-mediated pathways in myeloid leukemia cells. Mol. Cancer Ther. 4, 381–388.
Xu, Y.X., Pindolia, K.R., Janakiraman, N., Chapman, R.A., Gautam, S.C., 1997. Curcumin inhibits IL1 alpha and TNF-alpha induction of AP-1 and NF-kB DNA-binding activity in bone marrow stromal cells. Hematopathol. Mol. Hematol. 11, 49–62.
Zhao, W., Entschladen, F., Liu, H., Niggemann, B., Fang, Q., Zaenker, K.S., Han, R., 2003. Boswellic acid acetate induces differentiation and apoptosis in highly metastatic melanoma andfibrosarcoma cells. Cancer Detect Prev. 27, 67–75.