Dipartimento di Medicina Clinica e Sperimentale Direttore Prof. Mario Petrini
Dipartimento di Patologia Chirurgica, Medica, Molecolare e dell'Area Critica Direttore Prof. Paolo Miccoli
Dipartimento di Ricerca Traslazionale e delle Nuove Tecnologie in Medicina e Chirurgia
Direttore Prof. Giulio Guido
CORSO DI LAUREA SPECIALISTICA IN
PSICOLOGIA CLINICA E DELLA SALUTE
“Oscillazione lenta del sonno ed apprendimento visuo-motorio:
studio psicofisiologico mediante EEG ad alta densità”
RELATORE
Dott. Angelo Gemignani
CANDIDATO
Dott. Jacopo Agrimi
A nonno Ugo
In memoria del Professor Mario Guazzelli
RINGRAZIAMENTI
A Marco, Danilo, Francesca, Paolo, Lorenzo ed Andrea i componenti del gruppo Extreme, professionisti straordinari che lavorano con dedizione e coraggio in un paese dove la ricerca non riceve il valore che merita. Sono stati vicini a me in questi due anni non solo come insegnanti, ma anche come amici.
A Francesca, la mia compagna, per avermi sostenuto, capito ed aiutato in questi mesi di duro lavoro.
Ad Emanuela, Angelica e Mabel, i miei angeli custodi che hanno contribuito al lavoro di questa tesi.
Alla mia famiglia, non potevo desiderarne una migliore, che ha sempre appoggiato e sostenuto tutte le strade che ho scelto di percorrere.
RIASSUNTO
Background: Il concetto di “Oscillazione Lenta del Sonno” (“Sleep Slow Oscillation” –
SSO) è stato introdotto da Mircea Steriade (1993) mediante tecniche di registrazione intracellulare. La SSO è stata studiata attraverso approcci in computo ed in vivo sia nel modello animale che nell’uomo. Nell’uomo alcuni lavori hanno identificato le caratteristiche spazio-temporali dell'onda che corrispondono a specifici meccanismi molecolari: (i) picco negativo (downstate); (ii) picco positivo (upstate); (iii) Slope1 (sincronizzazione di apertura dei canali al potassio attività-dipendenti nel downstate); (iv)
Slope2 (sincronizzazione dell’attività di scarica dei neuroni all’uscita dal downstate); (v)
viaggio corticale, (Massimini et al. 2004, Menicucci et al. 2009, Piarulli et al. 2010). Recentemente alcuni lavori hanno posto l’attenzione su un'eccitazione (deflessione positiva) precedente al cosiddetto "silenzio elettrico" (downstate). Tale deflessione gioca un ruolo cruciale per tutta la dinamica dell'onda (esito e propagazione) in associazione con la "proneness" (massiva apertura dei canali al potassio in modo sincronizzato) alla bistabilità (Menicucci et al. 2013, Laurino et al. 2014). Oltre ad interessare alcune funzioni del sonno (downscaling sinaptico, incoscienza), la SSO rappresenta il comportamento cellulare e di network alla base del consolidamento delle memorie (Born e Rash, 2013). L’oscillazione lenta permette il replay della traccia mnestica inducendo interazioni cortico-ippocampali, mediate principalmente dall'attività dei fusi del sonno e dall’attività gamma sull’upstate. In sintesi, l’oscillazione lenta del sonno è un fenomeno complesso caratterizzato da origini, detezioni, caratteristiche morfologiche, dinamiche e da modulazioni di ritmi rapidi coalescenti con l’upstate. Queste caratteristiche rendono la SSO l’onda pivotale per i meccanismi dell’assenza di coscienza durante il sonno e per il consolidamento delle tracce mnestiche.
Scopo: lo scopo di questa tesi sperimentale è modulare la plasticità loco-regionale
mediante nuovi apprendimenti al fine di identificare le regioni “trigger” della SSO e di conseguenza anche il suo ruolo funzionale. In quest’ottica l’oscillazione lenta del sonno diventa “tout court” una finestra di osservazione elettiva per mappare i meccanismi di consolidamento dei nuovi engrammi mnestici durante il sonno.
apprendimento implicito di tipo visuo-motorio, ovvero un compito cinematico (Ghilardi et al. 2000). Lo studio è suddiviso in due parti di 24 ore, una di controllo (A) ed una sperimentale (B). L’esperimento, suddiviso in cinque punti salienti: ore 8, 13, 19, addormentamento e sonno, ore 8 del mattino successivo prevede per ogni punto in cui il soggetto è sveglio:
1) misura di ormoni correlati allo stress; 2) 15 minuti di EEG basale, 3) compito cinematico con (B) o senza (A) apprendimento implicito; 4) 15 minuti di EEG basale. Durante il sonno viene registrato l'EEG per l'intera durata della notte.
Risultati principali: (1) per quanto attiene le detezioni emerge che il compito mnestico
induce una differenza altamente significativa della quantità di eventi nelle aree premotorie e motorie nell’emisfero dominante;
(2) l’apprendimento del compito visuo-motorio si associa ad un incremento significativo
dei ritmi rapidi in banda sigma (12-16 Hz) ed in banda beta (16-25 Hz) che emergono sull’upstate, nelle aree di integrazione sensoriale retrorolandiche.
Conclusioni: (1) l'oscillazione lenta del sonno veicola il consolidamento delle memorie
acquisite durante la veglia nelle regioni coinvolte nelle singole dimensioni di un compito complesso, da quelle puramente motorie o di programmazione motoria a quelle di integrazione sensori-motoria. Questi risultati confermano che la SSO è l’onda pivotale per tutti i fenomeni di plasticità sinaptica LTP-dipendenti, che coinvolgono l’apprendimento. Ciò accade attraverso la coalescenza del fenomeno bistabile con ritmi rapidi in bande sigma e beta, di natura cortico-talamo-corticale, classicamente associate al fenomeno del replay della traccia mnestica;
(2) nel solco del lavoro di Laureys (2001) sul ruolo del REM nel consolidamento degli apprendimenti visuo-motori, i risultati di questa tesi portano ad ipotizzare, riprendendo la
sequential-step hypothesis di Giuditta del 1995, che il NREM e REM siano legati da
meccanismi comuni alla base della rimodulazione plastica degli apprendimenti Hebbiani.
Struttura della Tesi: per maggior leggibilità, la presente tesi è divisa in due parti, la
prima e principale, strutturata come un articolo scientifico, presenta all’inizio un’introduzione succinta, che partendo da una breve ricapitolazione della letteratura, affronta presto lo scopo della tesi, a cui seguono risultati e discussione. La seconda parte (dal cap. 5) invece fornisce una più estesa esposizione compilativa e dettagliata dei risultati in letteratura.
ABSTRACT
Background: The concept of Sleep Slow Oscillation (SSO) was introduced by Mircea
Steriade (1993) using intracellular recording techniques. SSO was studied adopting both
in computo and in vivo techniques, and both in animal models and in humans. In humans,
some works have identified the spatio-temporal characteristics of the wave, corresponding to specific molecular mechanisms: (i) negative peak (downstate); (ii) positive peak (upstate); (iii) Slope1 (potassium-channels opening synchronization); (iv) Slope2 (synchronization in firing activity of neuron exiting the downstate), and (v) cortical travel (Massimini et al. 2004, Menicucci et al. 2009 Piarulli et al. 2010). Some recent papers have focused on an excitation (positive deflction) preceding the so-called "electrical silence": this early peak is crucial to the whole dynamic of the wave (outcome and propagation) in association with the "proneness" (massive synchronized opening of potassium channels) to bistability (Menicucci et al. 2013, Laurino et al. 2014). Beside playing a role on some sleep functions (synaptic downscaling, unconsciousness), SSO represents the cellular and network mechanism at the basis of the consolidation of memories (Born and Rash, 2013). In fact, SSO allows the replay of the mnestic engram inducing cortico-hippocampal interactions, mostly mediated by the activity of sleep spindles and by gamma activity crowned upon the upstate.
In summary, SSO is a complex phenomenon characterized by sources, detections, morphologies, dynamic modulations of rapid rhythms coalescing with the upstate. All these features make the SSO wave pivotal to the mechanisms of the absence of consciousness during sleep and for memory consolidation.
Aim: The purpose of this thesis is to modulate the loco-regional plasticity via new
learnings, in order to determine regions triggering SSO and its functional role. In this perspective, the SSO becomes "tout court" the most important observation window to map the mechanisms of consolidation of new memories during sleep.
Methods: In line with Huber and co-workers (2004) we chose an implicit-learning task
of visual-motor type, i.e. a kinematic task (Ghilardi et al. 2000). The protocol is divided into two 24 hours’ modules, a control (A) and an experimental (B) one. Each module is divided into five main temporal points: 8 a.m., 1 p.m, 7 p.m, sleep, 8 a. m. of the
following morning. For each temporal point when the subject is awake the set of measures was:
1) stress-related hormones levels; 2) 15-minute baseline EEG, 3) computer-aided recording of a kinematic task with (B) or without (A) implicit learning; 4) 15-minute EEG. During sleep, the EEG was recorded for the entire duration of the night.
Main results: (1) Detections showed that the mnestic task induced a highly significant
difference in the amount of events in the motor and premotor areas in the dominant hemisphere; (2) The learning of visuo-motor task induced a significant increase of rapid rhythms in the sigma (12-16Hz) and beta (16-25 Hz) bands, crowning the upstate in sensory integrative regions behind the central sulcus.
Conclusions: (1) SSO conveys the consolidation of memories of vigil activity in the
regions involved in the single dimensions of a complex task, from the purely motor or motor-programs dimension, to that of sensory-motor integration. These results confirm that SSO is thus pivotal for all phenomena of LTP-dependent synaptic plasticity, involving learning. This occurs through the coalescence of the bistable phenomenon with rapid rhythms in the sigma and beta bands, whose cortico-thalamic-cortical nature is classically associated with memory traces replay. (2) Along the lines of the work of Laureys (2001) on the role of REM sleep on consolidation of visuo-motor learning, the results of this thesis lead to the hypothesis, reappraising “the sequential-step hypothesis” of (Giuditta et al. 1995), that NREM and REM sleep share common mechanisms underlying the plastic reshaping of Hebbian learning.
Structure of the Thesis: For readability, this thesis is divided into two parts, the first and
principal, structured as a scientific paper, moving from a succinct introduction, with a brief recapitulation of the literature, but fastly dealing with the scopes of the thesis, is followed by main results and a final discussion. The second part (from chapter 5) provides a more extensive and detailed compilatory exposition of already published results.
INDICE
1
INTRODUZIONE 102
MATERIALI E METODI 152.1 Partecipanti 15
2.2 Protocollo sperimentale 15
2.3 Registrazioni EEG e pre-elaborazione dei dati 16
2.4 Analisi dei dati 16
2.5 Compito visuo-motorio 17
3
RISULTATI 204
DISCUSSIONE 255
REVISIONE DELLA LETTERATURA:RAPPORTO TRA SLEEP SLOW OSCILLATION ED APPRENDIMENTO 27
5.1
SSO: Definizione e prime interpretazioni
27
5.2 Prime ipotesi funzionali: la Sleep Slow Oscillation
viaggia attraverso il mantello corticale
29
5.3 Studi su caratteristiche morfologiche e funzionali
della SSO nel 2009 e nel 2010
35
5.4 SSO, nuove ipotesi strutturali e funzionali
43
6 SONNO E APPRENDIMENTO
47
6.1 Storia di un’ipotesi
47
6.2 Riattivazione della traccia mnestica, NREM e REM
48
6.3 Funzioni della Slow Wave Activity nel
consolidamento delle memorie: l’ipotesi omeostatica
55
6.4 SWA ed apprendimento loco-regionale, Reto Huber
58
1 INTRODUZIONE
Nel 1993 un cambiamento di paradigma teorico-sperimentale ha caratterizzato la storia della fisiologia del sonno e delle sue funzioni: Mircea Steriade, Igor Timofeev e Florin Amzica identificarono che le fasi di sonno ipersincronizzato o profondo sono sostenute da uno specifico comportamento cellulare: tutti i neuroni corticali, siano essi eccitatori o inibitori, presentano un’oscillazione del potenziale di membrana, caratterizzato da fasi di silenzio elettrico (iperpolarizzazione profonda o downstate, tra -80 e -84 µV) alternate a fasi di attività di scarica (depolarizzazione o upstate) sovrapponibili in termini di coerenza spazio-temporale a quelle di comune riscontro nella veglia o nel sonno REM. Questo comportamento, definito da Mircea Steriade “Oscillazione Lenta” (<1Hz), rappresenta il fenomeno cellulare alla base di tutte le attività a bassa frequenza, come ad esempio le onde delta e i complessi K (risposta corticale ad uno stimolo sensoriale simile ad un’oscillazione tipica dello stadio 2 del sonno), (Loomis et al. 1937). Questi fenomeni contribuiscono in modo significativo alla determinazione della Slow Wave Activity (SWA), ovvero la frequenza principale (0.5-4Hz) che caratterizza il sonno NREM stadi 3 e 4 (detti anche Slow Wave Sleep - SWS) e più marginalmente lo stadio 2 del sonno NREM (Fig 1)
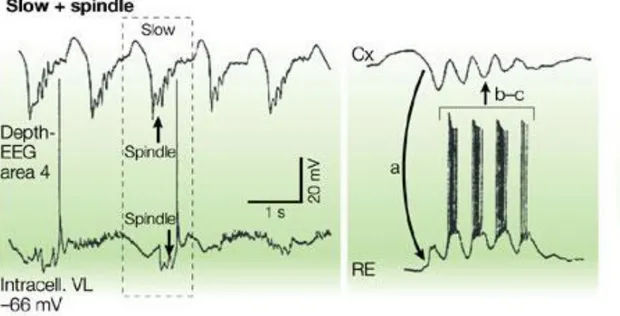
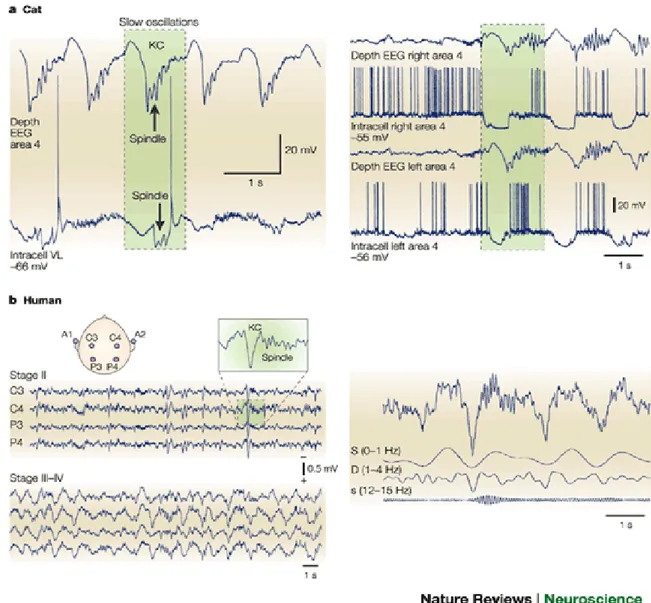
Fig. 1 Steriade 2004, Registrazioni intracellulari da neuroni corticali e talamici nel gatto sotto anestesia: le due immagini mostrano le oscillazioni lente ed i fusi del sonno
combinati tra loro, nel pannello di sinistra osserviamo una registrazione elettroencefalografica dei neuroni corticali dell’area 4 ed una registrazione intracellulare dai nuclei del talamo ventrolaterali. L’onda lenta negativa della frequenza circa di 0,9 Hz è seguita da una sequenza di fusi del sonno della frequenza di 10 Hz.
Una delle funzioni della SWA è quella di favorire il consolidamento notturno di engrammi mnestici acquisiti durante la veglia (Born e Rasch 2013). Nel 2004, Huber e collaboratori hanno identificato per la prima volta un ruolo cruciale della SWA nella memoria implicita, in particolare nei fenomeni di plasticità neurale associati ad apprendimenti di tipo visuo-motorio. In questo lavoro dall’analisi spettrale è emersa una relazione causale tra l’aumento della SWA nell’intervallo frequenziale della SSO e aree corticali coinvolte nell’integrazione visuo-motoria. Questi AA non hanno però affrontato il ruolo reale della SSO, sia in termini di forma e di topologia che di attività rapide associate all’upstate. Questa tesi si pone l’obiettivo di legare la struttura e la coalescenza con i ritmi rapidi tipica della SSO, con una topologia specifica compatibile con fenomeni plastici loco-regionali, fenomeni legati ad incrementi segregati di sinapsi Hebbiane durante la veglia. Come descritto fin quasi dalla sua scoperta, la SSO si caratterizza per una “coalescenza” con ritmi rapidi dall’attività sigma (12-16 Hz) tipica dei fusi del sonno fino all’attività gamma (superiore ai 30 Hz) tipica delle funzioni cognitive/emotive, (Steriade et al. 1993), (Massimini et al. 2004), (Tononi et al. 2006), (Gemignani et al. 2012), (Menicucci et al. 2009), (Born e Rasch 2013). L’ipotesi più affascinante relativa a questa coalescenza slow-fast è che i ritmi rapidi giochino un ruolo nell’indurre fenomeni di consolidamento delle tracce mnestiche (Sejnowski e Destexhe
2000). Recentemente, Menicucci e collaboratori, (2013) e Laurino e collaboratori, (2014) hanno mostrato una distribuzione temporale specifica nella espressione dei ritmi rapidi legati alla SSO. Grazie a questi lavori, il “dogma” della bifasicità (downstate-upstate) dell’oscillazione lenta è stato ampiamente superato, non solo in termini strutturali ma soprattutto funzionali e di viaggio corticale. L’oscillazione lenta è ad oggi un fenomeno trifasico caratterizzato da una prima deflessione positiva costituita da ritmi rapidi (definito “pre-downstate”) che precede il picco negativo (silenzio elettrico o downstate) che a sua volta promuove il successivo upstate (attività di scarica veglia-simile). La
domanda che si sono posti questi AA è quale sia il ruolo funzionale del “pre-downstate”. La risposta a questo quesito è stata sciolta grazie al modello sperimentale dei potenziali evocati durante il SWS. Come descritto in una review seminale di Amzica e Steriade (2000) i potenziali evocati durante il sonno o complessi K presentano gli stessi meccanismi cellulari delle SSO spontanee. Laurino e collaboratori (2014) hanno identificato che la prima risposta corticale ad uno stimolo sensoriale durante il sonno (sia esso acustico, tattile o visivo) avviene circa 200 msec dopo la presentazione dello stesso (P200) e presenta una topologia modalità-sensoriale specifica con latenze minori nelle rispettive aree sensoriali primarie (Fig 2). Per contro le latenze del downstate (N550) e del successivo upstate (P900) sono modalità-sensoriali indipendenti con una localizzazione aspecifica nelle regioni fronto-centrali. Laurino e collaboratori hanno interpretato questa risposta bistabile modalità-sensoriale indipendente (down+up-states) come specificamente cruciale per il mantenimento dell’incoscienza e quindi atta ad abolire ogni tentativo di elaborazione sensoriale. Nello stesso anno, Menicucci e collaboratori hanno confermato che ciò che è vero per le SSO evocate (complessi K) vale anche per quelle spontanee, ovvero che queste ultime presentano un bump positivo che precede il downstate (picco negativo). A questo punto rimane da chiarire il ruolo di innesco del pre-downstate o P200 nell’induzione del silenzio elettrico. L’ipotesi scientifica risiede nella identificazione e definizione da parte di Maria Sanchez-Vives dei canali al potassio-attività/depolarizzazione dipendenti (Sanchez-Vives et al. 2010). Per indurre bistabilità è necessario un surplus eccitatorio: se consideriamo il pre-downstate o la P200 alla stregua di eccitazioni corticali glutammatergiche allora possiamo concludere che è proprio la presenza di queste attività e il loro viaggio corticale (come qualsiasi eccitazione durante la veglia) ad innescare la bistabilità e tutti i meccanismi ad essa connessa come ad es. l’assenza di coscienza downstate-dipendente e i ritmi rapidi sull’upstate associati al consolidamento delle memorie. L’ultima domanda che dobbiamo porci è perché la bistabilità è maggiore nelle aree fronto-centrali. In altre parole perché queste regioni di natura associativa presentano una maggiore proneness alla bistabilità rispetto ad altre della stessa natura come ad esempio quelle del “planum temporale”. La risposta a questa osservazione risiede principalmente in due aspetti: 1) le strette relazioni che legano le regioni fronto-centrali e le circuiterie corticali distribuite alla base del mantenimento della coscienza;
2) una maggiore diffusione delle sinapsi Hebbiane in loco. Le aree fronto-centrali nell’uomo infatti giocano un ruolo sia di hub della coscienza (Sporns, 2013) che di regioni in cui avvengono in misura più abbondante la formazione di sinapsi Hebbiane LTP-dipendenti (Tononi e Cirelli 2003, 2006). Immaginiamo quindi che la P200, ad es. indotta da uno stimolo visivo, viaggi attraverso il dorsal ed il ventral stream verso le regioni anteriori incontrando passo dopo passo aree a diversa proneness per la bistabilità. Il risultato è che anteriormente la slope 1 (misura della caduta in down-state, Fig. 3) del complesso K è molto più ripida che in altre regioni per una maggiore presenza e sincronizzazione di apertura dei canali al potassio attività-dipendenti. Questo fenomeno molecolare fornisce la base per comprendere come usando come punto di “repere” il picco negativo, l’analisi del viaggio corticale metta in evidenza uno spostamento antero-posteriore del silenzio elettrico. Questo viaggio è solo apparente in quanto il viaggio reale è sostenuto da una eccitazione che precede la caduta in down-state. Lo stesso concetto vale per le SSO spontanee ed è sostenuto dal pre-downstate.
Emerge con chiarezza come la topologia delle latenze della P200 si modifichi in base al tipo di stimolo sensoriale, mentre quella del picco negativo (N550) e dell’upstate (P900) rimangano invariate. Latenze pari a 0 ms (evidenziate in blue scuro) appaiono nella prima riga, quella riferita alla P200, nelle aree temporali per gli stimoli acustici, nelle aree occipitali per gli stimoli visivi e nella corteccia somato-sensoriale per gli stimoli tattili.
Alla luce di queste osservazioni risulta facilmente comprensibile lo scopo di questa Tesi: modulare la plasticità loco-regionale mediante nuovi apprendimenti al fine di identificare le regioni trigger della SSO e del suo ruolo funzionale. In quest’ottica la SSO diventa “tout court” una finestra di osservazione elettiva per mappare i meccanismi di consolidamento delle nuove memorie durante il sonno. A questo riguardo, in linea con Huber e collaboratori (2004) è stato scelto come esercizio di apprendimento implicito di tipo visuo-motorio, un compito cinematico usato nei protocolli sperimentali di Ghilardi e collaboratori (2000).
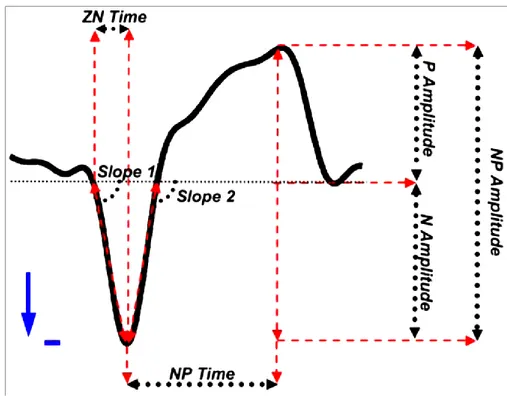
Fig 3 Piarulli 2010, è stata utilizzata questa figura per definire alcune caratteristiche morfologiche della SSO. Slope1: misura di sincronizzazione di apertura dei canali al potassio nel downstate. Slope2: misura della sincronizzazione dell’attività di scarica in uscita dal downstate.
2 MATERIALI E METODI 2.1 PARTECIPANTI
Fino ad oggi, sette volontari sani (2 femmine e 5 maschi, destrimani, tra i 20-27 anni) sono stati arruolati in questo studio. Nessun soggetto ha presentato in anamnesi una storia di disturbi psichiatrici, neurologici o internistici. Ogni soggetto ha trascorso una notte in laboratorio come adattamento alla condizione sperimentale. Ai soggetti è stato richiesto di evitare uso di alcol e caffeina nelle 24h che precedono il protocollo. I dati sono stati ottenuti lasciando al soggetto libertà di dormire al proprio orario di abitudine ed utilizzando il medesimo equipaggiamento per la registrazione EEG.
2.2 PROTOCOLLO SPERIMENTALE
Lo studio è stato suddiviso in due parti di 24 ore, una di controllo ed una sperimentale. I soggetti sono stati assegnati all’una o all’altra condizione in maniera randomizzata. L’esperimento prende avvio alle 8 di mattino ed è stato suddiviso in cinque punti salienti: alle ore 8, alle ore 13, alle ore 19, addormentamento e sonno (circa tra le 23 e le 24), ed infine alle ore 8 del mattino successivo. Ognuno di questi momenti è stato caratterizzato in sequenza da:
1) prelievo, tramite utilizzo di un tampone salivare di CORTISOLO, BDNF e NGF; 2) registrazione EEG, di 15 minuti, in veglia ad occhi chiusi;
3) compito cinematico usato nei protocolli sperimentali (Ghilardi et al. 2000), (Debarnot et al. 2009);
4) registrazione EEG, di 15 minuti, in veglia ad occhi chiusi;
Il momento sperimentale dell’addormentamento e sonno è invece l’unico che segue una struttura differente che consta dei soli punti 1 e 3 precedentemente descritti. E’ stato chiamato “giorno A” la condizione sperimentale di controllo, durante la quale il compito visuo-motorio non prevede la rotazione del cursore digitale (vedere 2.5) Invece, è stato chiamato “giorno B” la condizione sperimentale caratterizzata dalla suddetta rotazione del cursore. Dopo revisione della letteratura in merito all’utilizzo del compito cinematico
descritto in 2.5, sono stati utilizzati due angoli di deviazione della traiettoria del joystick: 15° e 45°. Il compito consta di 5 sessioni da 90 secondi ciascuna.
ORARIO DEVIAZIONE DEVIAZIONE DEVIAZIONE DEVIAZIONE DEVIAZIONE
8:00 0° 0° 15° 15° 15°
13:00 15° 15° 15° 45° 45°
19:00 45° 45° 45° 45° 45°
23:00 45° 45° 45° 45° 45°
8:00 45° 45° 45° 45° 45°
2.3 REGISTRAZIONI EEG E PRE-ELABORAZIONE DEI DATI
Le registrazioni EEG ad alta densità sono state eseguite usando un amplificatore Net Amps 300 (GSN300; Electrical Geodesic Inc., Inc., Eugene, OR, USA) e cuffie HydroCel Geodesic; il sistema comprende 128 elettrodi uniformemente distribuiti sullo scalpo. I dati sono stati acquisiti con una frequenza di campionamento di 500 Hz, una risoluzione di 24 bit, una precisione di 70 nV/bit e i segnali sono stati filtrati con una banda passante tra 0.01-500 Hz. L’elettrodo corrispondente al Vertex (CZ nel sistema di riferimento internazionale 10/20) è stato adottato come referenza di registrazione online. I valori di impedenza degli elettrodi sono stati mantenuti al di sotto dei 50 kOhm. Tutti i segnali sono stati registrati e visualizzati durante il sonno usando il software Net Station, versione 4.4.2 (Electrical Geodesic Inc., Inc., Eugene, OR, USA). Dopo l’acquisizione, i dati EEG grezzi sono stati importati e analizzati in MatLab (MathWorks, USA). Tutte le mappe sono state ottenute usando la tollbox di MatLab (EEGLAB). Durante l’analisi i dati EEG grezzi sono stati successivamente re-referenziati al potenziale medio valutato sugli elettrodi mastoidi, allo scopo di ottenere potenziali di tipo monopolare.
2.4 ANALISI DEI DATI
Gli stadi di sonno, veglia, N1 (stadio 1), N2 (stadio 2), N3 (stadi 3 e 4) e REM sono stati valutati in fase di analisi, mediante i criteri del AASM, tramite analisi visiva e spettrale di epoche di tracciato di 30 secondi. Le epoche contaminate da artefatti di natura non
elettrofisiologica (ad esempio movimenti artefattuali ) sono state eliminate. In ogni epoca priva di artefatti sono stati rilevati canali molto rumorosi usando una tecnica basata sull’analisi delle componenti principali (PCA). I canali rumorosi non sono stati considerati nell’analisi dell’epoca nella quale sono stati rilevati. Nelle epoche appartenenti agli stadi N2 ed N3 è stata effettuata la rilevazione automatica delle SSO mediante i criteri di detezione utilizzati da Piarulli e collaboratori (2010). Questo metodo prevede la detezione degli eventi SSO completi, ossia gli eventi con almeno un’onda che durante la propagazione raggiunge le soglie dei criteri di detezione. Il metodo inoltre permette la selezione di canali contenenti onde SSO sottosoglia che effettivamente completano la mappatura di ogni evento propagante. I criteri per la detezione sono i seguenti (Massimini 2004) : A) due zero crossings separati da 0,3-1 sec con il primo avente slope negativa; B) il picco negativo tra i due zero crossings con ampiezza negativa inferiore a -80 microvolts; C) un’ampiezza negativo/positivo di almeno 140 microvolts. Per ogni SSO individuata abbiamo stimato 4 caratteristiche morfologiche (Menicucci et al. 2009, Piarulli et al. 2010): Due caratteristiche di ampiezza (ampiezza picco negativo, ampiezza picco positivo), ed infine due caratteristiche riguardanti le slopes (la pendenza dal primo zero crossing al picco negativo e quella tra il picco negativo ed il secondo zero
crossing). Per ogni evento è stato considerato come canale di origine dello stesso quello
che presentava minore latenza del picco di downstate rispetto agli altri canali dello stesso evento. Inoltre la modulazione delle attività rapide (in banda sigma 12-16Hz, beta 16-25 Hz e gamma 25-35 Hz) è stata valutata come differenza tra la potenza spettrale, in tali bande, stimata nella fase di upstate rispetto a quella di downstate. Le potenze in upstate e
downstate sono state calcolate su finestre di Hamming di 500 msec centrate
rispettivamente nei picchi di upstate e downstate. La comparazioni delle caratteristiche delle SSO tra notte A e notte B sono state eseguite con un test non parametrico, test di Wilconxon. E’ stato preferito questo test a causa della bassa numerosità dei soggetti. Stessa logica ci porterà ad analizzare le correlazioni tra punteggi del compito visuo-motorio e features delle SSO con il test di Spearman con significatività al 5%.
2.5 COMPITO VISUO-MOTORIO
Passiamo ora in rassegna le caratteristiche dell’esercizio proposto ai soggetti secondo le prime interpretazioni di Ghilardi e collaboratori (2000) e secondo quelle più recenti,
avanzate e complesse di Debarnot e collaboratori (2009). Il compito di apprendimento visuo-motorio implica una maggiore plasticità sinaptica locoregionale molto specifica rispetto ad una condizione di controllo di non apprendimento. E’ stato scelto questo tipo di paradigma sperimentale, in linea a quanto pubblicato da Huber e collaboratori (2004) per l’estrema efficacia del compito, il suo essere esente da variabili non controllate e soprattutto per la sua capacità di elicitare l’attivazione di aree cerebrali specifiche e circoscritte. Tale specificità risulta utile per andare ad indagare gli effetti del sonno sull’apprendimento di tipo visuo-motorio. Ghilardi e collaboratori (2000), per esaminare l’attivazione corticale durante l’esecuzione e l’apprendimento di movimenti complessi, hanno utilizzato una famiglia di compiti motori cinematici e dinamici che chiamano in causa abilità esecutive, mnemoniche e cognitive ben differenziate. I soggetti che hanno preso parte al lavoro di ricerca di Ghilardi e collaboratori, dovevano raggiungere su di uno schermo degli obiettivi illuminati in sequenze semi-random muovendo un cursore digitale con la propria mano dominate (tutti i soggetti erano destrimani), a partire da una posizione centrale di partenza. Nel nostro lavoro si è ripetuta la stessa procedura, ma abbiamo deciso di privilegiare il joystick al cursore digitale. Durante la sessione sperimentale era presente un bias direzionale della traiettoria, la quale risultava ruotata di 15°-45°.
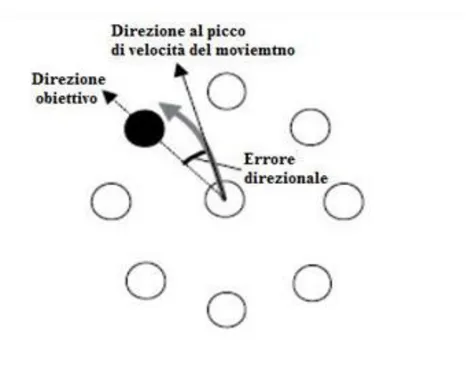
Fig 3 il compito visuo-motorio a cui sono sottoposti i soggetti. Si noti la rotazione del cursore. I soggetti muovono un joystick, eseguendo movimenti in avanti ed indietro da una posizione di
partenza centrale, puntando su uno degli otto obiettivi (distanza di 4.2 cm) visualizzati sullo schermo di un computer insieme alla posizione del cursore. Gli obiettivi si illuminavano in maniera random ad intervalli regolari di 1 secondo. Nel compito di adattamento alla rotazione, condizione non conosciuta dal soggetto, la posizione del cursore veniva ruotata in senso antiorario relativamente alla posizione della mano dei soggetti per mezzo di un angolo di deviazione fisso. In una sessione separata, almeno due settimane prima o dopo, il soggetto doveva ripetere la performance senza alcuna rotazione del cursore. L’errore direzionale prestabilito, è stato computato come l’angolo tra la linea della posizione iniziale della mano rispetto alla posizione dell’obiettivo (linea tratteggiata) e la linea della posizione della mano al momento del picco di velocità di movimento (linea solida). La linea grigia rappresenta la traiettoria della mano.
3 RISULTATI
I risultati principali, da sviluppare ulteriormente in futuro, di questo lavoro di Tesi indicano che le modificazioni plastiche delle circuiterie corticali, indotte dall’apprendimento del compito visuo-motorio implicito durante la veglia, portano durante il sonno ad un incremento significativo:
1) della “detezione” delle SSO nelle aree premotorie e motorie dell’emisfero dominante (sinistro), aree che giocano un ruolo nella programmazione motoria;
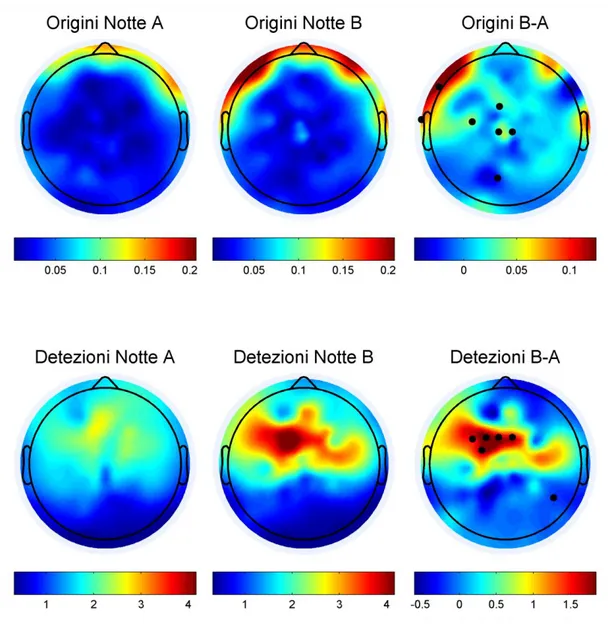
2) della modulazione sigma e beta sull’upstate topologicamente specifica: maggiore nelle aree posteriori al solco di Rolando che giocano un ruolo nella integrazione visuo-motoria. Per quanto riguarda la topologia delle origini alcuni canali mostrano differenze significative tra le due condizioni (giorno B vs giorno A), in posizione frontale e centrale (prossimi a Cz) (Fig. 4, prima riga).
Per quanto attiene le detezioni (Fig. 4, seconda riga) emerge una differenza altamente significativa della quantità di eventi nelle aree premotorie e motorie nell’emisfero dominante (sinistro). Non a caso tutti i soggetti erano destrimani.
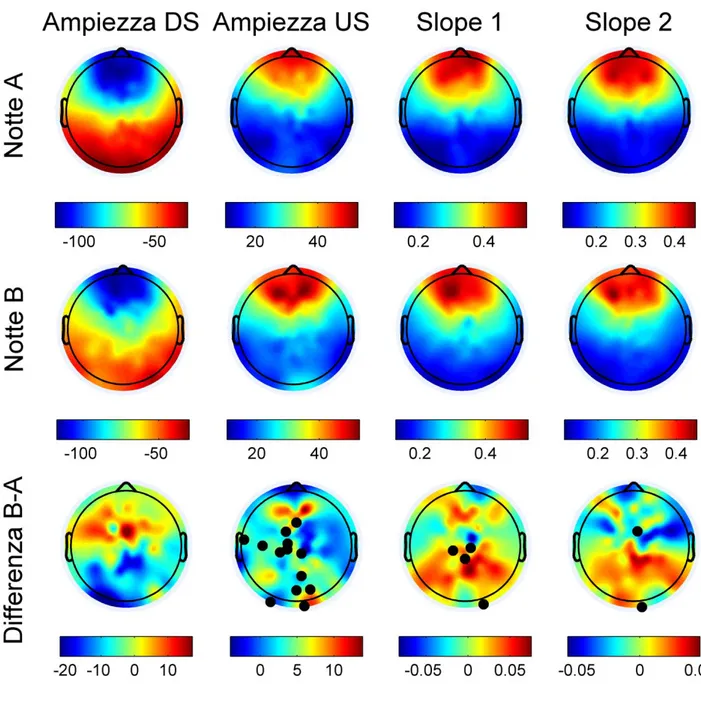
I successivi approfondimenti hanno affrontato l’analisi delle caratteristiche morfologiche delle SSO (Ampiezza dowstate, Ampiezza upstate, slope1, slope2). Come si può osservare nella Fig. 5, il giorno B induce una maggiore ampiezza dell’upstate nelle aree centrali, motorie sinistre e nelle aree posteriori di integrazione visiva.
Per quanto attiene la sincronizzazione di apertura dei canali al potassio nel downstate (slope1) e la sincronizzazione dell’attività di scarica in uscita dal downstate (slope2) si osservano punteggi significativamente più elevati nelle aree centrali (entrambe le slopes sono più ripide), Fig.5. Per completezza dell’analisi della morfologia si riporta anche il dato relativo al template delle SSO nelle due condizioni sperimentali. In Fig. 7 si riportano i templates medi per N2 e N3 relativi al giorno A e B.
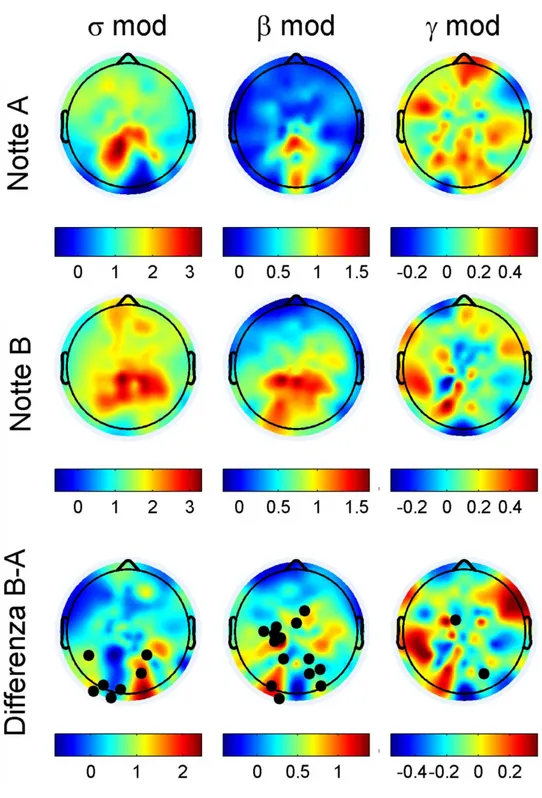
L’altro risultato principale di questo lavoro di Tesi è a carico della modulazione sigma e beta sull’upstate. L’apprendimento del compito visuo-motorio induce un incremento significativo dei ritmi rapidi in banda sigma (12-16Hz) ed in banda beta (16-25 Hz) che emergono sull’upstate, nelle aree di integrazione sensoriale retrorolandiche (che includono anche la corteccia visiva) Fig. 6.
Fig 4: Nella prima riga della figura è mostrata la topologia delle origini delle SSO. In ordine da sinistra verso destra: la topologia delle origini delle SSO nella notte A (controllo), notte B (condizione sperimentale), Sono riportati, in valore assoluto, il numero di eventi rilevati per minuto dal blu scuro (0 SSO) fino al rosso scuro (4 SSO). B-A è la differenza calcolata tra le due notti,i puntini neri rappresentano il singolo elettrodo caratterizzato da differenza significativa tra le due notti (p ≤ 0,05%). Nella seconda riga della figura sono mostrate le detezioni delle SSO in termini di singoli eventi per minuto da sinistra verso destra: nella notte A, B e la differenza fra le due. Emerge con chiarezza in rosso scuro e attraverso i singoli elettrodi segnalati da punti neri la differenza significativa fra le due condizioni.
Fig 5: Nella prima colonna è mostrata l’ampiezza del downstate (DS) nella notte di controllo (Notte A), nella condizione sperimentale (Notte B) e la differenza fra ambedue (Differenza B-A) espressa in Microvolts (µV). Nella seconda colonna è stata fatta la stessa analisi quantitativa, ma relativa all’ampiezza dell’upstate (US) sempre in (µV) . I puntini neri rappresentano il singolo elettrodo caratterizzato da differenza significativa tra le due notti (p ≤ 0,05%). Si noti la forte modulazione in positivo dell’ampiezza nelle aree motorie, pre-motorie e di integrazione visuo-motoria. Nella terza e nella quarta colonna troviamo i dati relativi alla slope 1 e slope 2 nelle due notti ed il delta tra le due notti espressa in µV/msec.
Fig 6 : modulazione delle attività rapide in banda sigma 12-16Hz, beta 16-25 Hz e gamma 25-35 Hz espressa in decibel. Nella prima riga la notte di controllo (Notte A), nella seconda la condizione sperimentale (Notte B), nella terza la differenza tra le due notti (B-A). I puntini neri rappresentano il singolo elettrodo caratterizzato da differenza significativa tra le due notti (p
≤ 0,05%). Si noti come in frequenza sigma e beta vi siano forti modulazioni positive nelle
Fig. 7 Template delle SSO in N2 nella colonna di sinistra ed in N3 in quella di destra. Nella riga superiore la notte A di controllo, nella riga inferiore la notte B della condizione sperimentale.
4 DISCUSSIONE
I risultati principali di questo lavoro di Tesi permettono di tracciare alcune conclusioni: 1. l’oscillazione lenta del sonno è un fenomeno complesso caratterizzato da origini,
detezioni, caratteristiche morfologiche, dinamiche e da modulazioni di ritmi rapidi coalescenti con l’upstate. Queste caratteristiche rendono la SSO l’onda pivotale per i meccanismi dell’assenza di coscienza durante il sonno e per il consolidamento delle tracce mnestiche.
a. La forte differenza di detezioni di eventi SSO per minuto nella condizione sperimentale rispetto ai controlli nelle aree pre-motorie e motorie (area 6 ed area 4 di Brodmann) si associa a meccanismi di plasticità che coinvolgono queste stesse aree durante l’apprendimento del compito. Le aree premotorie, area 6 di Broodmann (e parzialmente quelle motorie primarie, in particolare l’area 4b), sono implicate nei processi di integrazione di azioni complesse eseguite o immaginate (motor imagery). L’osservazione dell’incremento delle detezioni in queste regioni non è quindi un risultato inaspettato. A differenza di quanto osservato da Huber e collaboratori (2004), il compito cinematico impone al soggetto una serie complessa di azioni che vanno dal movimento puro al feedback sensomotorio e visivo, fino alla rielaborazione mentale dell’esercizio stesso (Buccino et al. 2007, Debarnot et al. 2009).
b. Per quanto attiene le modulazioni dei ritmi rapidi sull’upstate, l’aumento significativo in banda sigma 12-16 Hz ed in banda beta 16-25 Hz rappresenta l’espressione elettrofisiologica del replay della traccia mnesica che permette il consolidamento degli apprendimenti nelle aree di integrazione visuo-motoria. Questo concetto è in linea con quanto descritto da Mölle e collaboratori nel 2011 in studi sugli spindles. Il dato relativo ad una maggiore modulazione in banda sigma e beta spiega anche l’aumento di ampiezza dell’upstate sia nelle aree di integrazione visuo-motoria che in quelle di programmazione motoria. Questi dati sono in linea con il ruolo dei fusi del sonno (la cui frequenza è in banda sigma e anche parzialmente
in banda beta) nel replay delle nuove tracce mnestiche acquisite in veglia (Born e Rasch, 2013).
c. La maggiore sincronizzazione di apertura dei canali al potassio attività-dipendenti (slope1) nelle aree centrali ed anteriori conferma la maggiore
proneness di queste regioni alla bistabilità e quindi a legarsi in modo
aspecifico ai meccanismi LTP-dipendenti indotti dalla veglia. Questo aspetto assume un ruolo cruciale nell’evitare l’emergenza di pattern di attività integrati su larga scala modulati da sinapsi Hebbiane distribuite sul mantello corticale. D’altro canto l’assenza di coscienza permette ai micro moduli corticali di funzionare in modo segregato e favorire quindi l’ottimizzazione delle circuiterie corticali utili non solo per il consolidamento delle memorie ma anche, come ad esempio messo in evidenza in studi recenti, per la clearance della beta-amiloide (Xie et al. 2013) .
2. Un altro intrigante risvolto di questo lavoro riguarda la prospettiva di integrare il ruolo del sonno NREM nel consolidamento delle memorie con quello del REM nel solco della sequential-step hypothesis (Giuditta et al. 1995) a lungo dibattuta in letteratura. Il lavoro di Laureys e collaboratori del 2001 ha dimostrato tramite fMRI come un compito di memoria implicita visuo-motorio modulasse la connettività funzionale tra corteccia premotoria sinistra (nei soggetti destrimani) e la corteccia parietale posteriore sinistra ( area 7 di Brodmann) da un lato e dall’altro con l’area supplementare motoria (area 6 di Brodmann). Il sonno REM guiderebbe il replay della traccia mnestica acquisita in veglia. Nel nostro lavoro abbiamo trovato da un lato un forte incremento di detezioni di SSO nell’aree premotorie di sinistra e dall’altro un forte aumento della modulazione sigma/beta nelle aree associative parietali. Ci chiediamo allora se il fenomeno da noi osservato possa essere letto come un meccanismo sequenziale comune che coinvolga il NREM ed il REM nel consolidamento dei nuovi apprendimenti legati alle catene di programmazione delle azioni visuo-motorie. (Rizzolatti 2007).
5 REVISIONE DELLA LETTERATURA: RAPPORTO TRA SLEEP SLOW OSCILLATION ED APPRENDIMENTO
5.1 SSO: Definizione e prime interpretazioni
Il sonno è un fenomeno fisiologico dinamico e complesso, cruciale per l'omeostasi dell'intero organismo. Nel suo lavoro del 1993, tramite registrazioni intracellulari in vivo nel gatto, Mircea Steriade ha mostrato che le fasi di sonno NREM sono sostenute da uno specifico comportamento cellulare. Tutti i neuroni corticali, siano essi eccitatori o inibitori presentano un’oscillazione del potenziale di membrana, caratterizzato da fasi di silenzio elettrico (iperpolarizzazione o downstate) alternate a fasi di attività di scarica (depolarizzazione o upstate) sovrapponibili in termini di coerenza spazio-temporale a quelle di classico riscontro nella veglia o nel sonno REM. Questo comportamento, definito da Mircea Steriade “Oscillazione Lenta” (<1Hz), rappresenta il fenomeno cellulare alla base di tutte le attività lente del sonno, come ad esempio le onde delta ed i complessi K, che contribuiscono in modo significativo alla determinazione della Slow Wave Activity, ovvero la frequenza principale (0.5-4Hz) che caratterizza le fasi del sonno NREM stadio 2 e in particolare del sonno NREM stadi 3 e 4 (detti anche Slow Wave Sleep - SWS). La SSO è caratterizzata da una prima fase di profonda iperpolarizzazione (downstate) durante la quale i neuroni corticali sono virtualmente silenti, che è seguita da depolarizzazione della membrana (upstate). (Steriade et al. 1994), (Steriade e al. 2001), (Steriade et al. 2003a ), (Steriade et al. 2003b ). Sia il downstate che l'upstate costano di alcune centinaia di millisecondi, durante l'upstate l'intero sistema talamo-corticale è investito da un'intensa attività sinaptica. L'oscillazione lenta è innescata, mantenuta e terminata attraverso il contributo di correnti intrinseche e network integrati talamo-corticali Fig. 8 (Timofeev et al.2000), (Timofeev et al. 1996 A) (Timofeev et al. 1996 b).
Solo recentemente grazie ai lavori di Giulio Tononi da una parte e di Jan Born dall’altra si è cominciato a far luce sul ruolo della SWA, rimasto fino a quel momento estremamente nebuloso. Tononi, riparafrasando Feinberg (1974), afferma che l’attività lenta del sonno è legata in maniera omeostatica alla necessità di dormire, ed in modo originale descrive come il suo andamento aumenti con la veglia e diminuisca nel corso
della notte. La funzione di questo complesso processo sarebbe la “potatura” e il riordino delle connessioni in eccesso createsi durante il giorno risultanti in un’accresciuta forza sinaptica. Questo fenomeno di potatura interessa in modo globale ed aspecifico tutto il
cervello. Born è invece convinto che l’attività lenta del sonno guidi un processo ippocampale di riattivazione delle tracce mnestiche acquisite in veglia, favorendone così il consolidamento.
Fig. 8 Steriade 2004 a) registrazioni intracellulari durante la fase NREM del sonno nel gatto, b) registrazione EEG nell’uomo (stadio III e IV ) sonno NREM..
5.2 Prime ipotesi funzionali: la Sleep Slow Oscillation viaggia attraverso il
mantello corticale
Si deve al nostro paese un notevole passo in avanti nell’interpretazione del fenomeno SSO. Il lavoro di Marcello Massimini del 2004 rappresenta infatti una pietra angolare negli studi sull’attività lenta in continuità con i risultati di Steriade. Nello studio del 2004 Massimi si interroga sulla natura del fenomeno SSO: se si tratti di un’onda globalmente sincrona o se al contrario viaggi attraverso la corteccia, con un preciso sito di origine e un altrettanto preciso percorso.
Il primo risultato che emerse, come dalle attese, fu che la SSO non comparisse né nello stato di addormentamento, né durante il REM, sporadici episodi potevano essere riscontrati in stadio 2, sotto forma di complessi K. Al contrario con l'approfondirsi del sonno in stadio 3 e 4 il fenomeno aumentava decisamente. Dalla fig. 9 tratta dal lavoro di Massimini emerge con chiarezza che la SSO, seppur caratterizzata da una distribuzione globale sullo scalpo, è rintracciabile in maniera nettamente più marcata nelle regioni frontali e parietali. Come emerge dalla figura 9 (C e D) gli elettrodi in cui è maggiore la possibilità di rintracciare l'oscillazione lenta sono, secondo la media, quelli anteriori corrispondenti ai lobi frontali. Al contrario i canali dove vi è la minima possibilità sono ubicati nelle aree temporali e occipitali. La parte più intrigante del lavoro riguardava però lo studio delle latenze tra l'occorrenza dell'onda nei vari elettrodi. In particolare il tempo di occorrenza della comparsa del picco negativo venne rilevato elettrodo per elettrodo e sovrapposto. Il risultato emerge in maniera chiara dalla figura 10 (A) in cui sovrapponendo i segnali provenienti da tutti gli elettrodi in un singolo ciclo di SSO emerge lo shift dovuto ai diversi tempi di occorrenza. La figura 10 (B) illustra la disposizione spaziale dei ritardi tra i picchi negativi del medesimo ciclo di SSO a partire dall'origine indicata dall'asterisco rosso. Si osserva che il gradiente di tali ritardi aumenta con progressione regolare dalle aree frontali sinistre alle parietali destre. Questi fecero ipotizzare come la SSO non fosse un evento locale e globalmente sincrono, ma come piuttosto viaggiasse attraverso lo scalpo seguendo percorsi riproducibili a
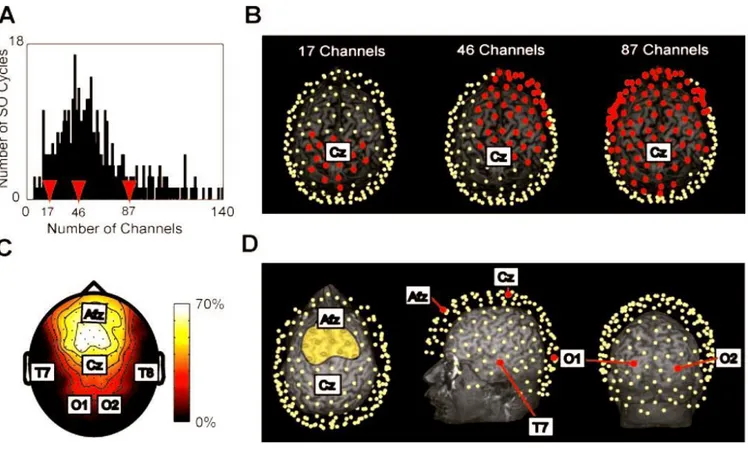
Fig. 9 Massimini et al. 2004 Ogni oscillazione lenta influenza diversi sottoinsiemi di canali.
"A" mostra la distribuzione del numero dei canali investiti da un ciclo di oscillazione lenta che si presenta durante un'ora di sonno di un soggetto. La maggior parte dei cicli è stato riscontrato circa in 50 elettrodi. Le frecce rosse indicano il numero dei canali influenzati dai tre cicli di oscillazione lenta rappresentati in "B", dove la posizione degli elettrodi sollecitati (punti rossi) è mostrata rispetto alla MRI individuale in 3D. Da notarsi che ogni onda agisce su un sottoinsieme di canali che variano sia in numero che in posizione. "C", la “detection density map” illustra la probabilità di individuare l'oscillazione lenta. Quasi il 70% delle oscillazioni lente sono state individuate nelle regioni fronto-centrali, mentre gli elettrodi parietali ed occipitali hanno rilevato poche o nessuna oscillazione lenta. in "D", le posizioni sullo scalpo dei sistemi di 10-20 elettrodi rappresentati sulla mappa in 2D di "C" sono proiettati sulla MRI in 3D dello stesso soggetto. L'area gialla nella figura più a sinistra include gli elettrodi con la più alta probabilità di rilevare l'oscillazione lenta. Le proiezioni corticali di questi elettrodi corrispondono alle aree 6, 8 e 9. Le cortecce temporale ed occipitale hanno una bassa probabilità di detezione (<10%)
Fig.10 Massimini et al. 2004, ogni ciclo di oscillazione lenta si propaga come una “traveling wave.”
"A" mostra i segnali registrati dai canali investiti da un singolo ciclo di oscillazione lenta disposti dal punto più alto (top) al punto più basso (bottom) secondo il ritardo del picco negativo. Da notare che l'oscillazione lenta non è precisamente sincronizzata in tutti i canali e che può essere misurata una continua distribuzione di intervalli di tempo. La larghezza dell'area rossa rappresenta il ritardo massimo (120 msec) dal picco negativo dal punto top del tracciato, al picco negativo al punto bottom del tracciato. "B" raffigura la distribuzione spaziale dei ritardi sulla “delay map”. Un asterisco rosso segnala la posizione del canale con ritardo pari a zero (l'origine). Le linee blu che partono dall'origine rappresentano le streamlines calcolate sul campo del vettore dei ritardi. L'oscillazione lenta origina localmente e si propaga ordinatamente al resto dello scalpo come una “traveling wave”. In "C" gli stessi segnali di "A" sono sovrapposti.
Sempre i dati dello stesso lavoro testimoniavano come successivi cicli di SSO occorressero in siti di origine differenti e con diversi patterns di diffusione.
Tuttavia una mappa precisa delle streamlines dell'onda e dei punti di origine di quest'ultima mostrava come gli elettrodi anteriori fossero investiti con una frequenza assai maggiore rispetto ai temporali e agli occipitali e come la direzione dell'onda fosse con più probabilità fronto-occipitale. In particolare i siti di origine risultavano essere collocati nell'area di transizione tra la corteccia frontale dorsolaterale e orbitale. Fig 11 A,B,C,D
Fig 11 Massimini et al. 2004, le oscillazioni lente originano più frequentemente nelle regioni anteriori e si propagano in una direzione antero-posteriore.
“A” espone la mappa delle streamlines dalla prima ora di sonno in un soggetto (le oscillazioni lente che influenzano meno di 20 canali sono escluse). La grandezza di ogni punto è direttamente proporzionale al numero dei cicli originati da ogni elettrodo. Da notarsi che virtualmente è possibile qualsiasi schema di origine e di propagazione, sebbene gli elettrodi anteriori tendano a dar luogo a più oscillazioni lente e le streamlines che si spostano nella direzione antero-posteriore siano più numerose. In “B” è segnalata la probabilità di ogni elettrodo di essere l’origine di un’oscillazione lenta per ottenere una mappa di densità di origine. I punti con un’alta densità di origine sono stati individuati nelle regioni anteriori dello scalpo. La mappa del ritardo medio mostrata in “C” riflette la direzione anteroposteriore prevalente della propagazione dell’oscillazione lenta. “D” rivela che gli elettrodi con la più alta probabilità di essere l’origine si trovano in una regione dello scalpo situata al di sopra della transizione fra la corteccia dorso-laterale e l’orbitofrontale.
Fig 12 Massimini et al. 2004 pattern generale dell’origine e della propagazione dell’oscillazione lenta del sonno è riproducibile attraverso le notti e attraverso i soggetti.
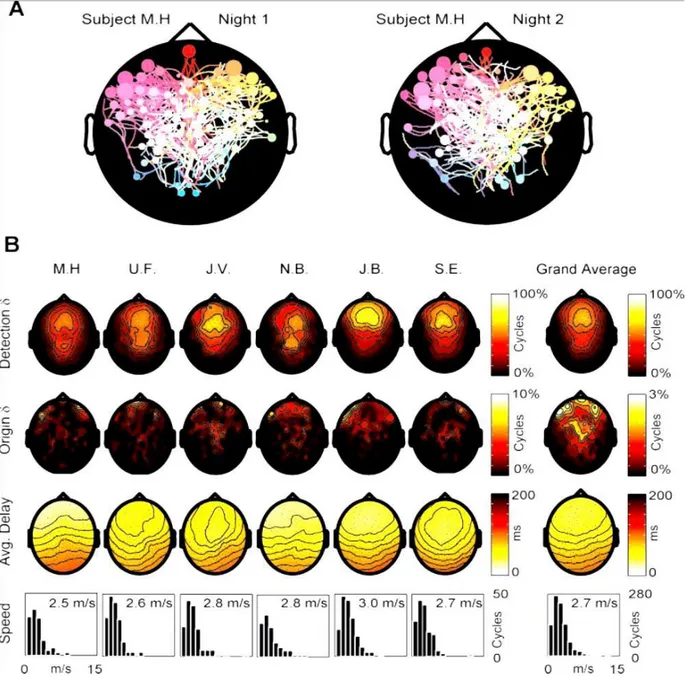
In “A”, le origini e i patterns di propagazione dei cicli di oscillazione lenta, registrate nello stesso soggetto durante due notti diverse, sono rappresentate da due mappe a streamlines. “B” riassume i risultati ottenuti da tutti i sei soggetti durante una notte. La prima fila contiene le mappe della densità delle detezioni che mostrano la probabilità di ogni elettrodo di rilevare l’oscillazione lenta. In tutti i soggetti, la più alta densità era sopra il lobo frontale. In due soggetti (U.F. e N.B.), un punto secondario di alta densità è stato rilevato su una regione dello scalpo più posteriore. La media generale delle densità di detezione è mostrata sulla sinistra. Da notarsi che in tutti i soggetti la densità delle origini è più alta nella regione frontale. Come
esposto nella media generale di sinistra, i gruppi di elettrodi con la più alta densità di origine si trovano sulle regioni più anteriori dello scalpo, situati al di sopra della corteccia orbitofrontale. La terza fila contiene le mappe delle medie dei “delays”. In tutti i soggetti la direzione prevalente di propagazione è l’anteroposteriore. La quarta fila illustra la distribuzione della velocità di propagazione e della velocità media calcolata per tutti i cicli che si muovono sull’asse anteroposteriore in ogni soggetto. La velocità media dell’oscillazione lenta è simile in tutti i soggetti (circa 2,7 m/s).
Il pattern generale sopra descritto di origine e di propagazione dell'onda era, secondo i dati di Massimini, riproducibile sia attraverso le notti che attraverso i soggetti (Fig 12). Precedenti registrazioni intracellulari effettuate in animali anestetizzati mostravano una sostanziale mancanza di sincronizzazione delle SSO sulla corteccia (Amzica e Steriade 1995a), (Amzica e Steriade 1995b). Il lavoro di Massimini spiegava tale desincronizzazione non come occorrenza casuale delle SSO bensì come una propagazione ordinata dell'onda attraverso lo scalpo. Risultava ancora ignoto il meccanismo alla base della determinazione del sito di origine dell'onda al cominciare di ogni nuovo ciclo, ma era plausibile ipotizzare che le sorgenti del fenomeno fossero collocate laddove durante il giorno si era accumulata maggiore forza sinaptica.
Studi in vitro suggeriscono che l'upstate di tale attività potrebbe provenire da neuroni dello strato 5 caratterizzati da firing spontaneo.
In più secondo Steriade la propagazione della SSO sarebbe mediata da connessioni cortico-corticali come suggerito dal fatto che l'onda sussiste anche in presenza di resezione del talamo e al contrario è interrotta dalla disconnessione chirurgica o farmacologica delle connessioni intracorticali.
A sostegno della tesi di Massimini secondo cui il sito d'origine della SSO sarebbe eminentemente frontale vi sono studi sul flusso sanguigno che indicano che quest'ultimo sarebbe molto più basso proprio nelle regioni frontali. (Braun et al. 1997)
Altri studi ancora mostrano come a seguito di deprivazione di sonno aumenti l'attività lenta nelle aree anteriori, (Finelli e al. 2000). Pare interessante riportare anche gli studi di Horne (1993) secondo cui la totale deprivazione di sonno indurrebbe sintomi molto simili a quelli di pazienti con lesioni orbito-frontali.
5.3 Studi su caratteristiche morfologiche e funzionali della SSO nel 2009 e nel
2010.
Il lavoro del centro Extreme della Scuola Superiore Sant’Anna di Pisa si è sforzato di approfondire caratteristiche morfologiche e funzioni dell'onda lenta sia come fenomeno a sé stante sia attraverso le sue relazioni con gli stadi del sonno nella sua globalità. Partendo dai primi lavori del 2009 si è giunti a conclusioni recentissime che in parte stravolgono la tesi stessa di Massini sulla propagazione dell’onda
I primi risultati del gruppo avevano confermato l'ipotesi secondo la quale ogni singolo evento di iperpolarizzazione originasse da un sito preciso per poi viaggiare lungo lo scalpo e che tali origini si ritrovassero con maggiore probabilità nelle aree frontali, come mostrato in Fig 13
Fig 13 Menicucci et al. 2009 La mappa di posizione dell’ origine e la distribuzione dell’estensione di propagazione.
A) Viene mostrata la media generale della mappa di densità di origine calcolata in base alle mappe individuali. Essa indica la frequenza (riportata in percentuale) dell'origine dell'episodio di SSO per ogni singolo elettrodo
La velocità media di propagazione dell'onda si assestava sul valore medio di 5 m/s, senza essere influenzata da altri fattori esterni. (Menicucci et al. 2009), (Piarulli et al 2010) Elementi assai interessanti emergevano dallo studio sull'ampiezza della propagazione di ogni singola onda. Durante lo stadio 2 del NREM infatti tale ampiezza risulta essere significativamente maggiore rispetto a quella registrata durante SWS (slow waves sleep). Inoltre si osservò come le SSO originate dalle aree occipitali viaggiassero di più rispetto a quelle originate nelle frontali. Il gruppo di ricerca aveva voluto testare quanto l'effetto della propagazione incidesse sulla distribuzione spaziale di detezione delle onde, distribuzione spaziale che aveva tra l'altro confermato i dati di Massimini sulla prevalenza frontale. Dai dati si poteva osservare che l'ampiezza di propagazione per area specifica fosse nettamente maggiore in posizione frontale e centrale rispetto alla posteriore come conseguenza della più alta quantità di SSO originate anteriormente, ma anche per la capacità di quelle occipitali di viaggiare attraverso lo scalpo. Infatti l'ampiezza di propagazione per specifica area è dipendente dal sito di origine di ogni SSO: gli eventi con origine frontale rimangono circoscritti a tale area, quelli occipitali invece vengono riscontrati in maggioranza nelle aree centrali, ma anche seppur in minor rilevanza nelle anteriori come mostrato in Fig 14 (Menicucci et al. 2009). Ci si chiedeva se la divisione dell'encefalo in due emisferi influenzasse la propagazione delle SSO. Dai dati del lavoro di Menicucci e collaboratori emergeva che il 75% degli eventi di SSO viaggiasse nell'emisfero controlaterale. Come conferma di ciò quasi tutti gli eventi che rimangono unilaterali sono quelli localizzati in un minor numero di elettrodi, quelli quindi che si propagano di meno. Il gruppo di Gemignani e collaboratori, dopo la propagazione, ha concentrato i suoi sforzi nella descrizione delle caratteristiche morfologiche degli eventi di SSO. La ricerca si è focalizzata su 7 aspetti ( features) : l'ampiezza è stata scomposta in tre misure, da un lato l'ampiezza generale dall'altro quella specifica del picco negativo e del picco positivo. Poi si è passati ad analizzare la slope, marcando come primo indice il valore assoluto della pendenza della curva tra il primo passaggio dallo zero al picco negativo, come secondo indice il passaggio dal picco negativo allo zero dell'upstate. Infine si sono misurati due indici di tempo, uno dal picco positivo al negativo, l'altro dal primo passaggio allo zero al picco negativo. La fig. 15 mostra perfettamente queste features che risultano cruciali non solo per la morfologia del fenomeno, ma anche per il suo ruolo funzionale.
Fig 14 Menicucci et al. 2009, ampiezza di propagazione nell'area come funzione della posizione di origine e degli stadi del sonno.
In ogni colonna sono mostrati gli eventi originati in un’area per mezzo della relativa mappa. Sulla sinistra sono raffigurate le mappe di detezione (% di eventi che interessa ogni elettrodo); sulla destra sono mostrati i corrispondenti contrasti fra le aree dell’ampiezza di propagazione nella regione.
Fig. 15 (Piarulli 2009) Definizione grafica delle caratteristiche morfologiche estratte.
Ogni onda individuata è stata selezionata per mezzo di sette caratteristiche: tre misure di ampiezza (l’ampiezza dal picco negativo al picco positivo -NP amplitude- , l’ampiezza del picco negativo -N amplitude - , l’ampiezza del picco positivo -P amplitude-), due misure di pendenza (il valore assoluto della slope fra il primo passaggio dopo lo zero e il picco negativo -slope 1- e fra il picco negativo e il secondo passaggio dopo lo zero -slope 2-), e due diverse stime di ampiezza temporale (il tempo dal picco negativo al picco positivo -NP time- e l’intervallo dal primo passaggio allo zero al picco negativo -ZN time-).
Passando all'analisi dei dati veri e propri ricavati dai soggetti, riportiamo la tabella dei valori medi riferiti a ogni singola caratteristica morfologica. ( Table 1) L'interesse del gruppo di ricerca si era per prima cosa focalizzato su come modificazioni delle features potessero intervenire nei differenti stadi del sonno. Merita riportare il fatto che in questo
caso le differenze tra i singoli soggetti hanno dimostrato di avere un ruolo significativamente rilevante. Elementi di grande interesse si rilevano nell'interazione, che risulta significativa all'analisi della varianza, tra gli stadi del sonno, l'origine dei singoli eventi e le caratteristiche morfologiche di quest'ultimi. (Table 2).
Dai dati riportati notiamo come alcuni parametri dei singoli eventi di SSO risultino significativamente più ampli in SWS rispetto allo stadio 2 del NREM a cui sono paragonati. Questo vale infatti sia per l'ampiezza dell'onda che per la durata della transizione dal down-state all'up-state.
Si noti anche dalla terza colonna che il sito d'origine di un ciclo di SSO influenza significativamente i valori inerenti alla slope. Questi dati sono ben esemplificati anche dalla fig 1.9 in cui viene mostrato il rapporto tra le caratteristiche morfologiche e il sito d’origine dell’evento. Ulteriori risultati sono stati ricavati dalla ricerca di una possibile interazione tra le caratteristiche morfologiche di un evento di SSO e il suo sito di detezione, ossia l'area sullo scalpo in cui questo veniva rintracciato. Come ben esemplificato dai dati della tabella l'area di detezione porta a dei mutamenti nelle features dei cicli di SSO.
Le SSO rintracciate nelle regioni frontali hanno la maggior ampiezza e anche il maggior tempo di durata, chiaramente in maniera indipendente dal sito di origine. Al contrario gli eventi rintracciati in corteccia temporale ed occipitale sono i più ridotti, sia per tempo che per ampiezza.
Il lavoro del gruppo di Gemignani focalizzò l'attenzione nella discussione finale del proprio articolo su come i dati sopra-riportati modificassero la nostra visione sulla SSO. Lo studio di Massimini veniva confermato nella tesi secondo la quale un singolo ciclo di SSO non fosse un fenomeno globalmente sincrono, ma al contrario si trattasse di un'onda che viaggia attraverso lo scalpo da un sito d'origine ben preciso. Era confermato anche il dato secondo cui vi sarebbe una maggiore possibilità di rilevare eventi di SSO nelle aree frontali piuttosto che nelle temporali e occipitali. Dai dati dello studio emersero però elementi prima non considerati che modificarono la visione del fenomeno. Si è rilevato che la pendenza (slope1) dell'onda nel passare dallo zero alla fase di down-state è maggiore in tutti i soggetti rispetto alla pendenza della stessa nella fase di passaggio all'up-state (slope2). Questo dato sembrerebbe confermare il lavoro di Volgushev e collaboratori che avevano già rilevato questa caratteristica in registrazioni intracellulari nel gatto. Quindi vi sarebbe maggior sincronizzazione del pool neuronale ingaggiato nell'evento nella fase di silenzio elettrico piuttosto che in quella di attività di scarica. Il cuore dello studio si era inoltre concentrato sull'interazione che lega sia il fenomeno di propagazione dell'onda che le sue caratteristiche morfologiche ad altre variabili quali gli stadi del sonno e il sito d'origine e di detezione dell'episodio SSO rilevato.
Nel lavoro prodotto da Massimini e Ferrandelli nel 2005 si è dimostrato che l'onda lenta indotta da stimolazione magnetica transcranica (TMS) durante il sonno profondo tende a non propagarsi e a rimanere circoscritta. Anche dai dati di Gemignani e collaboratori del 2009 emergeva che l'evento di SSO fosse più facilmente elicitabile in SWS piuttosto che in stadio 2, ma in SWS tendesse a non propagarsi attraverso lo scalpo e a rimanere nell'area in cui è originato. La tendenza alla nascita di un maggior numero di episodi di SSO, tra l'altro caratterizzati da parametri quali ampiezza e pendenza maggiore, durante lo SWS rispetto allo stadio 2 del NREM potrebbe essere dovuta ad un sempre maggior abbassamento, con l'approfondirsi del sonno, del tono di firing della formazione reticolare e dei nuclei colinergici mesopontini.
Fig 16 Menicucci 2009, Le caratteristiche morfologiche come funzione dell’origine.
Per ogni caraterisitica morfologica (feature) che mostra un' effetto sul luogo di origine, come riportato nella tabella 2, sono mostrati i contrasti fra le aree di origine. Ogni fila corrisponde ad una feature. Nella colonna di sinistra sono raffigurate le mappe delle medie generali delle features. Poiché le mappe corrispondono all'effetto sul luogo di origine, gli eventi che compaiono in ogni sito di origine (elettrodo) sono stati selezionati ed è stata derivata la media generale per ognuno degli stessi. Nella colonna destra osserviamo i risultati dei contrasti riportati graficamente con una scala di grigi. Ogni livello nella scala dei grigi indica una differenza significativa fra i toni contigui. La scala adottata è ordinale, cioè i toni più chiari corrispondono a valori relativi molto più alti per la features.
Tale meccanismo abbasserebbe il flusso eccitatorio talamo-corticale favorendo un più alto grado di iperpolarizzazione delle popolazioni neuronali. La difficoltà degli eventi di SSO a propagarsi durante SWS sarebbe invece causato dall'interruzione di ampli patterns di connettività cortico-corticale.
Probabilmente, esattamente come i complessi K in stadio 2, anche gli eventi di SSO durante SWS tendono ad inibire le risposte agli stimoli esterni, aiutando il soggetto a mantenere profondo il proprio sonno.
Dai dati di Menicucci e Piarulli veniva riconfermata la antero-posteriore come direzione prevalente di propagazione degli eventi di SSO. Si deve considerare plausibile l'esistenza di una via mesiale che ingaggerebbe il cingolo anteriore e posteriore e il praecuneo, (Murphi et al. 2009).
Elemento intrigante a cui riporre attenzione è la conferma dai dati del fatto che le aree frontali siano più portate a originare fenomeni di iperpolarizzazione di grandi popolazioni neuronali. Le motivazioni di ciò possono essere molteplici, sicuramente non sfugge il fatto che la frontale è un'area eteromodale massimamente attiva durante la veglia e portata quindi ad accumulare una gran quantità di informazioni e memorie.
Questa visione del fenomeno SSO, come descritta nei lavori del 2009 e del 2010, è stata profondamente rivoluzionata dagli studi di Gemignani e collaboratori del 2012 e da quelli del 2013 di Laurino e di Menicucci. Nel capitolo seguente verrà esposto “lo stato dell’arte” sull’oscillazione lenta del sonno come conseguenza delle ultime scoperte sul campo.
5.4 SSO, nuove ipotesi strutturali e funzionali.
Negli ultimi due anni sono usciti lavori che hanno gettato una nuova luce sulla struttura e la funzione dell’oscillazione lenta. Partiamo dallo studio su un caso singolo di una rara malattia genetica, l’insonnia fatale familiare, patologia che porta alla neurodegenerazione delle strutture talamiche, (Gemignani et al. 2012).
Se Steriade aveva correttamente dimostrato che l’iperpolarizzazione della SSO fosse un fenomeno di natura cortico-corticale, da questo studio emerge però come sia il talamo a giocare un ruolo cruciale nell’Upstate della SSO, Fig. 17.