TAPHONOMIC ANALYSIS ON FOSSIL REMAINS FROM THE CIOTA CIARA CAVE (PIEDMONT, ITALY) AND NEW EVIDENCE OF CAVE BEAR AND WOLF
EXPLOITATION WITH SIMPLE QUARTZ FLAKES BY NEANDERTHAL
FABIO BUCCHERI1, DAVIDE FEDERICO BERTÈ1,2*, GABRIELE LUIGI FRANCESCO BERRUTI2,3, ISABEL CÁCERES4,5, LISA VOLPE6 & MARTA ARZARELLO1
1*Corresponding author. Dipartimento di Studi Umanistici, Università degli Studi di Ferrara, C.so Ercole I d’Este 32. 44121 Ferrara, Italy. E-mail: [email protected]; [email protected]; [email protected]
2Associazione culturale 3P (Progetto Preistoria Piemonte), via lunga 38, San Mauro to.se, Torino, Italy. E-mail: [email protected] 3International Doctorate In Quaternary and Prehistory, Department of Geology, Universidade de Tras-os-Montes e Alto Douro, Quinta de Prados, 5000-801 Vila Real, (Portugal). E-mail: [email protected]
4Area de Prehistoria, Universitat Rovira i Virgili (URV), Avinguda de Catalunya 35, 43002 Tarragona, Spain.
5IPHES, Institut Català de Paleoecologia Humana i Evolució Social, Zona Educacional 4 - Campus Sescelades URV (Edifici W3) 43007 Tarragona, Spain. E-mail: [email protected]
6TekneHub and Dipartimento di Fisica e Scienze della Terra, Università degli Studi di Ferrara, Via Saragat,1. 44122 Ferrara, Italy. E-mail: [email protected]
To cite this article: Buccheri F., Bertè D.F., Berruti G.L.F., Cáceres I., Volpe L. & Arzarello M. (2016) - Taphonomic analysis on fossil remains from the Ciota Ciara Cave (Piedmont, Italy) and new evidence of cave bear and wolf exploitation with simple quartz flakes by Neanderthal. Riv. It. Paleont. Strat. 122(3): 41-54.
Abstract. The Ciota Ciara cave is located in the karst area of Monte Fenera (Borgosesia - VC) and, with the Ciotarun cave, it is the only Middle Palaeolithic site in Piedmont where the presence of Homo neanderthalensis has been confirmed by discoveries of human remains. Preliminary taphonomic and archaeozoological studies have been performed on a portion of the palaeontological remains from the Stratigraphic Unit 14 (1144 bones). The studies confirmed the presence of cut-marks on Ursus spelaeus and Canis lupus, made by lithic instruments. The po-sition of the cut-marks on the bones can be related to skinning and butchery. An experimental butchery has been performed to test the efficiency of the tools made by local quartz during slaughtering activities. The archaeozoo-logical analysis of the faunal remains of S.U. 14, identified cut-marks with weak peculiarities, probably due to the use of quartz tools. The analysis of the experimental collection allowed distinguishing between cut-marks made by quartz tools from those made by flint tools. A preliminary experimentation, conducted on more than 50 different cut-marks made with flakes of three different raw materials (vein quartz, quartzite and flint), allow us to hypothesize that it is possible to distinguish cut-marks made with unretouched flakes of different raw materials.
Received:March 10, 2016; accepted: September 20, 2016
Keywords: Homo neanderthalensis, Ursus spelaeus, archaeozoology, quartz, cut-marks, Middle Paleolithic.
I
ntroductIonThe relationship between Homo neanderthalen-sis and Ursus spelaeus has been debated for a long
time by researchers: until the first half of the XX century the abundant accumulation of cave bear bones in many sites was interpreted as a proof of Neanderthal men’s activity (see d’Errico & Giacobi-ni 1986 and citations therein). Koby (1951) recogGiacobi-ni- recogni-sed in natural processes the taphonomic origin of these bone accumulations in the European caves, and after Kurtén (1958) the use of the cave by be-ars for the mere purpose of winter hibernation was widely accepted. The general interpretation was that humans and cave bears were seasonally separated (de Lumley 1972). However, in some sites cave, bear
exploitation has been effectively demonstrated: i.e., Erd (Hungary; Gabori-Csank 1968), Scladina Cave (Belgium; Abrams et al. 2014) and Nietoperzowa Cave (Poland; Wojtal 2007). In other sites, evidence of use of brown bear has been found: Biache-Saint-Vaast (France; Auguste 2002), Fate Cave (Italy; Va-lensi & Psathi 2004), Fumane Cave (Italy; Jéquier et al. 2012).
At the Ciota Ciara Cave (Borgosesia, Vc, Piedmont, Italy, Fig. 1) cut-marks have been found on some carnivores’ bones, reopening the problem of human-carnivore interaction during the Pleisto-cene. During the archaeozoological analysis of 14 faunal remains from the Stratigraphic Unit 14 (S.U. 14), a number of cut-marks, showing some pecu-liarities, have been identified. We hypothesise that this could be related to the raw material used, i.e. vein quartz (Fernández-Jalvo & Cáceres 2010). To
confirm this hypothesis an experiment was carried out, performing butchering activities on animal car-casses using simple flakes of different raw material. One objective of this research is the iden-tification of possible morphological distinctions between cut-marks visible on bone surfaces produ-ced by unretouched lithic tools made with different raw materials. This research was inspired also by the preliminary works on the differentiation of the cut-marks conducted by Fernández-Jalvo and Cáceres
(Fernández-Jalvo & Cáceres 2010). The raw mate-rials considered in this study are flint, vein quartz and quartzite. The choice of these raw materials is due to their wide use throughout the whole prehi-story (e.g.: Lemorini et al. 2014; Daffara et al. 2014; Clemente & Gibaja 2009; Colonge & Mourre 2009; Lombera Hermida 2008; Brugal & Raposo 1999; Mourre 1996) and to the great differences in the edges morphology between vein quartz, quartzite, and flint tools.
Fig. 1 - The Ciota Ciara Cave; a: location map of Monte Fenera (Piedmont, Italy) and the Ciota Ciara Cave; b: map of the excavation site; c: stratigraphy of the Nord section (modified from Angelucci et al. 2015).
The site
The Ciota Ciara cave is located at 670 m a.s.l. in the karst complex of Monte Fenera (Borgosesia, Vercelli) (Fig. 1). The cave opens on the western side of the mountain. With other caves of the mount (i.e. Ciotarun cave), it represents one of the most im-portant and complete evidence of the Palaeolithic of Piedmont. Since 2009, the University of Ferra-ra, in collaboration with Soprintendenza Archeologia del Piemonte, has carried out excavation campaigns, in
the atrial area of the cave, that have led to the iden-tification of five main S.U.s with faunal remains and lithic industries: namely 13, 14, 15, 103 and 140 S.U. (Angelucci et al. 2015) (Fig. 1c). The lithic assem-blage is made of flakes, few retouched tools, cores, hammers and débris. Raw materials exploitation was
achieved through the direct percussion technique and through various methods: S.S.D.A. (Système par Surface de Débitage Alterné, Forestier 1993), discoid
and Levallois (Boëda 1988, 1993; Daffara et al. 2014). The small number of retouched tools reve-als a further adaptation to the characteristics of the raw material: the retouch on quartz flakes is quite difficult and it does not permit to obtain stronger or more useful edges in comparison to the unre-touched ones (Mourre 1996). Among the reunre-touched tools, most of them are side-scrapers, lateral or
con-vergent, followed by denticulates and notches (Daf-fara et al. 2014). Several raw materials are represen-ted in different proportion: quartz is predominant, followed by spongolite, sandstone, mylonite, opal and allochthonous flint of high quality (Arzarello et al. 2012; Berruto 2011; Daffara et al. 2014). The archaeological record consists of various typologies of quartz: macro-crystalline pegmatite quartz, mi-cro-crystalline pegmatite quartz and hyaline quartz. All these types of raw materials have been found, in secondary position in the proximity of the archaeo-logical site, within 5 km range (Arzarello et al. 2013; Daffara et al. 2014). In the Ciota Ciara cave, 27 to-ols made with allochthonous red flint of unknown provenance were found (Arnaud et al. 2014). This red flint is different from the grey flint that can be found in nodules included in carbonate levels from 720 m a.s.l. to the top of Mount Fenera (Berruto 2011). Tools obtained from the local flint are not represented in the archaeological record of the Cio-ta Cara cave.
The faunal assemblage is dominated by Ursus spelaeus, occurring with many other species of
mam-mals (Berto et al. 2016) (Tab. 1). The small mammal association from the Ciota Ciara cave (S.U. 13, 103 and 14) gives us information about the environment that surrounded the site and made it possible to establish the chronology of the site in the tempe-rate-humid period of MIS 5 (80-70 ka) (Arnaud et al. 2014). The environment was characterized by deciduous woodland and, probably at the base of the mountain, by glades (Berto et al. 2016). The in-tersection between different habitats, as well as the presence of lithic raw materials and water sources, were the main factors that favoured the human oc-cupation during the Late Pleistocene. The study of the faunal remains demonstrates a marked predo-minance of U. spelaeus in all the five stratigraphic
units. The occurrence of several cubs of U. spelaeus
suggests a recurring use of the cave as den (Berto et al. 2016).
Morphological variability of the cut-marks
Cut-marks are the result of butchering acti-vities (Shipman & Rose 1983) and, therefore, a ubi-quitous sign on the surfaces of the bones that have been processed for carcasses exploitation. The im-portance of cut-marks has been evident since the late 19th century (Lartet & Christy 1875) and their
Foglio1
Taxon NISP MNI NISP MNI NISP MNI
Ursus spelaeus 33 5 232 9 989 18 Ursus arctos 15 1 24 2 17 2 Panthera leo 1 1 14 2 Panthera pardus 5 1 Lynx lynx 1 1 4 1 Canis lupus 1 1 8 1 11 2 Vulpes vulpes 2 2 3 1 Meles meles 6 1 2 1 Martes martes 1 1 Rupicapra rupicapra 3 1 9 2 26 2 Cervus elaphus 8 2 104 5 cf. Dama 1 1 Bos primigenius 1 1 1 1
Bos vel Bison 1 1 1 1
Stephanorhinus sp. 4 2 Sus scrofa 1 1 Marmota marmota 68 3 Erinaceus europaeus 3 1 1 1 SU 103 SU 13 SU 14 Pagina 1
Tab. 1 - Faunal remains in the Ciota Ciara Cave subdivided for Strati-graphic Unit. NISP: number of individual specimens; MNI: minimum number of individuals.
relation with slaughtering activities was highlighted by Martin during his studies on the French Mou-sterian site “la Quina” (Martin 1909, 1910), but it is only during the second half of the 20th century that specific studies on cut-marks began (Shipman 1986). Cut-marks can be identified by considering their re-lative location on the bone, due to the butchering activity (flesh removal, fur removal, disarticulation and evisceration) and their morphology, due to the tools involved (i.e. stone tools, bamboo tools, iron tools etc.). All butchering activities (flesh removal, fur removal, disarticulation and evisceration) leave different kind and patterns of marks on different skeletal elements and bone regions (Binford 1981; Nilssen 2000).
Concerning the morphology of the cut-mar-ks, particular care has to be applied, since not only different tools can leave different marks, but also unretouched flakes produce morphologically diffe-rent cut-marks depending on the activity (Moretti et al. 2015).
Shipman and Rose’s research (Shipman & Rose 1983) allowed the clear distinction of cut-mar-ks from all the other taphonomic alterations, but they did not distinguish the cut-marks morphology depending on the raw materials (obsidian, lava, flint, basalt and quartzite) used during the experimenta-tion. Cut-marks made through flint flakes are well known in the archaeological contexts and are stu-died in detail by experimental archaeology (Walker & Long 1977; Shipman 1983; Galán & Domínguez-Rodrigo 2013), while a similar analysis on the diffe-rent lithic raw materials involved in the butchering process is still lacking. Many studies focus on the identification of possible morphological distin-ctions of the cut-marks visible on the bones surface (e.g. Olsen & Shipman 1988; Schick & Toth 1993; Giacobini 1995; Greenfield 1999, 2006; Choi & Driwantoro 2007; West & Louys 2007; Bello & So-ligo 2008; Bello et al. 2009; Domínguez-Rodrigo et al. 2009; de Juana et al. 2010; Yravedra et al. 2010). Several studies analyse the micromorphology of the cut-marks with the aim of differentiating the types of tools used to produce them; the most part focu-ses on the distinction between cut-marks produced with stone tools and cut-marks produced with va-rious types of metal tools. These studies were usual-ly carried out with optical microscopy and scanning electron microscopy (Olsen & Shipman 1988; An-drews 1990) and, more recently, through
three-di-mensional microscopes (Bello & Soligo 2008; Bello et al. 2011). Different studies demonstrated that there are morphological differences between cut-marks produced by metal or stone knives (Walker & Long 1977; Greenfield 1999), bamboo knives (West & Louys 2007), shell tools (Choi & Driwan-toro 2007), and lithic handaxes (Bello et al. 2009; de Juana et al. 2010). These researches stated that the morphology of the edge of the tool is a factor that influences the characteristics of the cut-marks on the bone.
The most important factor that determines the edge morphology of a lithic tool is the raw ma-terial. Several use wear and technological studies show that lithic tools made with different lithic raw materials have unretouched edges with diffe-rent characteristics (Lemorini et al. 2014; Daffara et al. 2014; Berruti & Arzarello 2012; Igreja 2009; Clemente & Gibaja 2009; Colonge & Mourre 2009; Lombera Hermida 2008; Mourre 1996).
MaterIalsandMethods Archaeozoological study
The archaeozoological study of the faunal remains of the Ciota Ciara Cave, was conducted on 1144 bones from S.U. 14, corre-sponding to the total of the faunal remains recorded during the exca-vation campaign in 2013 (Tab. 2), 36% of the entire bone assemblage of the S.U. 14. This faunal sample comes from an excavation area Tab. 2 - Faunal remains in the Ciota Ciara Cave founded from SU14
in 2013. NR: number of remains. Sheet1 Species NR % Ursus spelaeus 257 22.47 Ursus arctos 2 0.17 Canis lupus 7 0.61 Panthera leo 1 0.09 Lynx lynx 2 0.17 Martes martes 1 0.09 Mustelidae 1 0.09 Carnivora 1 0.09 Cervus elaphus 14 1.22 Bos primigenius 1 0.09 Rupicapra rupicapra 11 0.96 Cervidae 3 0.26 cf. Dama 1 0.09 Marmota marmota 40 3.50 Aves 1 0.09 Undet 801 70.02 TOTAL 1144 100.00 Page 1
of 8 square meters (F2, F3, F4, E3, E4, D2, D3, D4) placed in the atrium of the cave. The identifiable remains are subdivided in three groups according to the size of the mammals: large, medium and small. Mammal size groups are modified from Bunn et al. (1980) and Bunn (1986): 1) Very small size (<20 kg); 2) small size (20-100 kg); 3) medium size (100-300 kg); 4) large size (300-1000 kg); 5) very large size (> 1000 Kg). In the large size group were placed: adult Ursus spelaeus, Ursus arctos, Panthera leo, Cervus elaphus, Stephanorinus sp. and Bos primigenius. In the medium size group were placed: young Ursus spelaeus, Canis lupus, Lynx lynx, Vulpes vulpes, Sus scrofa, adult Rupicapra rupicapra and cf. Dama. In the small size group were placed: Martes martes, young Rupicapra rupicapra, and Marmota marmota.
On the sample were calculated the Minimum Number of Elements (MNE), or the minimum number of skeletal portion ne-cessary to account for observed specimens, and the Minimal Animal Unit (MAU), obtained dividing MNE by the number of times the element occurs in a complete skeleton of the same animal.
The archaeozoological analysis of the archaeological sam-ple was conducted in accordance with the literature (e.g. López-González et al. 2006; Marín-Arroyo et al. 2014; Cáceres et al. 2012; Fernández-Jalvo & Andrews 2003). The faunal remains of the Ciota Ciara cave are characterized by a frequent presence of post deposi-tional alterations (Buccheri 2014; Angelucci et al. 2015), like trample marks, cracks, hydric abrasion, carnivores intervention, limestone concretions, deposition of manganese oxide and roots alterations. The identification of trampling marks is based on microscopic crite-ria described in Domínguez-Rodrigo et al. (2009). The deposition of manganese oxide is visible on the 99% of the remains, with different degrees of intensity and coverage. For this reason, the study of the deposition of manganese oxide was conducted according to López-González et al. (2006). The archaeozoological analysis of the faunal remains was carried out using four different stereomicroscopes: Carl Zeiss (10x / 20), Leica MZ8, Leica EZ4D, and Leica S6D with ca-mera; one digital microscope, a Dinolight Am413T, and a Scanning Electron microscope SEM ZEISS EVO MA15-HR equipped by software OXFORD SmartMap EDS INCA Energy 250 X-Act for EDS chemical microanalysis (working set up: electron source LaB6 cathode, accelerating voltage 20 kV, variable pressure).
Experimental protocol
An experimental protocol was developed to verify the na-ture of the cut-marks. The experimental protocol has to be calibrated using the same raw materials and the same tools identified in the archaeological contexts (Moretti et al. 2015). The experimental pro-tocol performed is devised to standardise all the operations and to minimize the variables of the process. The experimental butchery of a wild boar and of an adult rabbit was performed by two expert experimenters using tools made with three different raw materials: flint, vein quartz and quartzite.
The choice of vein quartz and flint was made in accordance with the raw materials used in the Ciota Ciara cave, and the choice of quartzite was made to provide a control parameter in the experiment. Vein quartz was collected in two different areas and in two different forms: pebbles and prismatic blocks. The first set of pebbles was col-lected in the creek that flows at the base of Monte Fenera (Vercelli, Piedmont), less than 5 km from the Ciota Ciara cave, from the same area whence, according to the studies conducted, came quartz peb-bles used by the Neanderthals (Arzarello et al. 2012; Berruto 2011; Daffara et al. 2014). The second set of fine-grained quartzite pebbles was collected in the fluvial terrace of the Tejo river in Portugal.
The flint pebbles come from the Gargano promontory in the south of Italy. The use of this excellent raw material is documented in several Palaeolithic sites (e.g. the Paglicci cave and Pirro Nord; Arzarello et al. 2007; Palma di Cesnola 2006). This raw material was
chosen for the granulometric similarity with the allochthonous flint (of unknown provenance) found in the Ciota Ciara cave.
The prismatic blocks were collected in the Sessera Valley where were found some flakes made in local vein quartz that testify to a prehistoric frequentation of this area (Berruti et al. 2016).
All the tools used during the experiment are unretouched flakes obtained through the S.S.D.A. method because it was the most employed in the Ciota Ciara cave and because there are just a few re-touched tools (Angelucci et al. 2015; Arzarello et al. 2012; Daffara et al. 2014). In the experiment, lithic tools have been used on different anatomical portions of the carcasses in order to avoid interpretative errors (e.g. flint on right hind limb, quartzite on left hind limb, quartz on forelimb). Every action with every tool was marked to reconstruct the relation between the cut-marks and the type of lithic tool.
The wild boar carcass underwent a complete butchering pro-cess, whereas the rabbit carcass was already skinned, so the only ac-tion undertaken during the experimentaac-tion was the flesh removal.
After the experimental butchery, the bones were boiled until complete removal of the remaining periosteum and flesh. After this pro-cedure, the bones were placed in a bath of alcohol (50%) and distilled water (50%) for 48 hours, and then washed with distilled water in an ultrasonic cleaner for 3 minutes. Microscopic analysis of the bones was conducted. The considered criteria, modified from Dominguez-Rodrigo et al. 2009, for the description of the cut-marks are: slope of the edge, bottom section of the groove, internal streaking, type of trait, morphol-ogy of the terminations, shoulder effect and presence of herzian cones. The slope of the edge could be abrupt, half-abrupt or variable; the bottom section of the groove could be V shaped or U shaped; the internal streaking could be thin or wide; the trait could be simple, mul-tiple or variable; the morphology of the termination could be Y-shaped or simple; the shoulder effect could be present or absent; the herzian cones could be present or absent.
r
esultsThe experimental activities, conducted by two different operators, allowed to produce 56 different cut-marks on 20 different bones: 12 rabbit bones and 8 wild boar bones. The analysis was carried out considering various features to describe and to recognize the cut-marks. The slo-pe of the edge is abrupt in flint cut-marks, half-abrupt in those by quartz and variable in those by quartzite. The bottom section of the groove left by flint is acute while the ones left by quartz and quartzite are obtuse. Flint leaves narrow internal striations, while quartz and quartzite leave wide internal striation. The shoulder effect is more common in quartz and quartzite cut-marks than in those from flint. The morphology of the cut-marks termination could be Y-shaped both for flint and quartz. The herzian cones are present in cut-marks made by flint and absent in those made by quartz and quartzite.
The skeletal representation of all taxa is low except in Ursus spelaeus (Tab. 3). The MNE
and MAU evidences that appendicular elements are more abundant in the sample under study; the distal part of limb bones (metapodial, car-pal/tarsals and phalanx) are the more abundant elements.
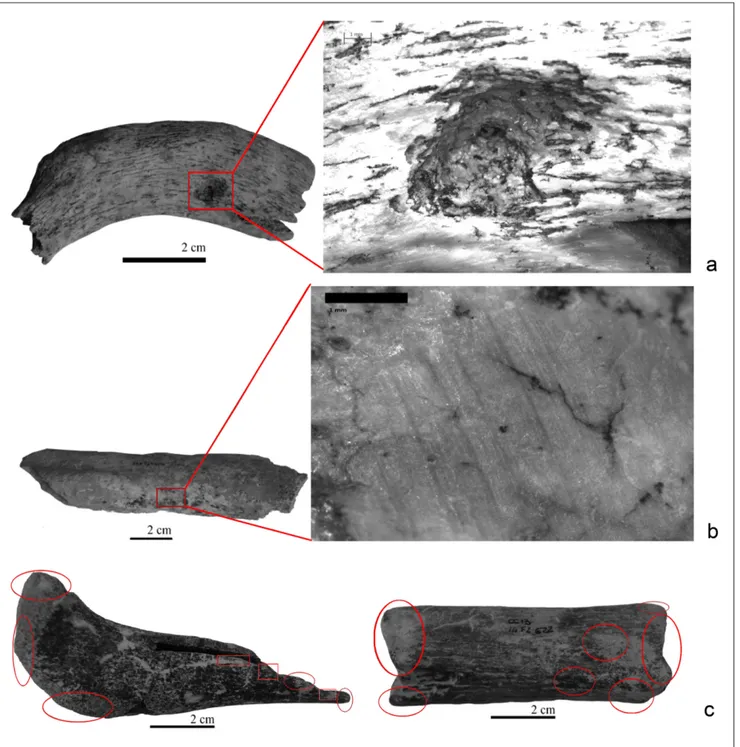
The study of the 1144 faunal remains has highlighted nine types of taphonomic alterations (Fig. 2): roots activities, cracking, carnivore inter-vention, deposition of manganese oxide, concre-tions, trampling marks, water abrasion (rounding and smoothing) and cut-marks (Tab. 4). Even so, in general, the taphonomical preservation of fos-sil assemblage is good.
Root marks are present on 730 remains, 63.81% of the sample examined; root marks that are white massive vermiculated trace, are located
on both the surface and on fracture edges, and do not deeply penetrate the bones. Carnivores inter-vention was recognized on 54.81% of the sam-ple; many bites are located on the fractured edge and on diaphysis without epiphysis; U. spelaeus is
the species with more traces of carnivore inter-ventions, equally reported on axial and appendi-cular skeleton. Cracking alteration was identified on 50.52% of the sample, to various degrees. Deposition of manganese oxide is detectable on 99.83% of the remains with different degrees of intensity and coverage; the manganese oxide pig-mentation is present in a generalized, dispersed or massive form without any type of standardi-zation, linked with the form, the size, the posi-tion and the taxonomy of the remains. The same Sheet1
MNE MAU MNE MAU MNE MAU MNE MAU MNE MAU MNE MAU
Antler 3 1.5 Skull 8 8 1 1 Hemimandible 3 1.5 Atlas 1 1 Axis 1 1 Cervical vertebra 1 0.2 2 0.4 Thoracic vertebra 3 0.25 Lumbar vertebra 3 0.42 Rib 4 0.17 Scapula 3 1.5 Homer 5 2.5 1 0.5 Radius 6 3 Ulna 3 1.5 1 0.5 Hamate 1 0.5 Scaphoid 2 1 1 0.5 1 0.5 Scapholunar 5 2.5 Trapezium 2 1 Trapezoid 1 0.5 I mc 4 2 II mc 4 2 III mc 3 1.5 IV mc 1 0.5 Pelvis 1 1 1 1 2 2 Femur 9 4.5 Patella 1 0.5 1 0.5 Tibia 3 1.5 Fibula 2 1 Talus 7 3.5 Calcaneus 2 1 1 0.5 Cuboid 1 0.5 Cubonavicular 1 0.5 I cuneiform 2 1 II cuneiform 1 0.5 III cuneiform 2 1 I mt 1 0.5 II mt 3 1.5 IV mt 2 1 Metapodial 5 0.25 1 0.25 2 0.5 I phalanx 21 1.05 1 0.13 II phalanx 15 0.75 2 0.25 1 0.13 III phalanx 12 0.6 1 0.13 Bos primigenius Anatomical element Ursus
spelaeus Ursus arctos Canis lupus elaphusCervus Rupicapra rupicapra
Page 1
Tab. 3 - Anatomical elements in the sample. MNE: minimum number of elements; MAU: minimal animal unit.
kind of distribution, without a clear pattern, cha-racterizes also the concretions (22.46% of the faunal remains). Trample marks are present on 30.90% of the remains; trampling traces are ran-domly distributed on bones, without prevalence for dimension of the fragments. Water alterations are represented by smoothing and rounding on 28.49% of the considered remains; the hydric alteration is prevalent on fragments with length between one and four cm and with elevated de-gree of fossilization. Thirtyfive different bones,
2.97% of the sample analysed, show on the sur-faces evidence of butchering. Twelve of them have been identified on taxonomy determinable bones of two different species: one Canis lupus
and eleven U. spelaeus remains (Fig. 3). Half of the
remains with cut-marks are undeterminable, but the other half are attributable to large or medium-sized animals (Tab. 5). The archaeozoological study of the cut-marks has allowed the identification of four dif-ferent activities: flesh removal, evisceration, fur re-moval and disarticulation (Nilssen 2000) (Tab. 6). Fig. 2 - Taphonomic alterations; a: bite marks; b: trampling marks; c: hydric abrasion.
The cut mark on C. lupus is located on the
third cervical vertebra. The cut-marks on U. spelaeus
are: on a skull fragment; on the atlas; on a thoracic vertebra; on a proximal part of a rib (Fig. 4); on two right humeri; on a right radius; on two left tibias; on a right fibula; on a right scapholunar.
d
IscussIonThe experimental activities confirmed the presence of small differences and peculiarities be-tween the cut-marks made with simple flakes of different raw material (Fernández-Jalvo & Cáceres 2010) (Tab. 7). The unretouched tools, used by the two operators for the same action, are differentiated only by their raw materials that generate cut-marks with different features (Fig. 5). These data suggest that the morphology of the cut-marks is linked with the raw material. The difference is probably due to the texture of the raw material that drives the char-acteristics of the functional edges (Fig. 6).
The texture of the raw material influences the possibility to obtain better functional edges, there-fore amorphous rocks, are the best kind of raw ma-terial. Flint is an amorphous rock, so no preferential fracture plans are present, thus allowing to knap flint in a predetermined way and to obtain very sharp edges (Arzarello et al. 2011). Conversely, vein quartz is made by crystals. Its structure consists of a trigonal crystal system, and these characteristics made vein quartz more difficult to knap in order to obtain tools (Mourre 1996; Arzarello et al. 2011). The edge of a quartz tool is less strong than one made in flint, but very sharp. Quartzite is a meta-Foglio1 Size NR % NR % NR % NR % Big 5 14.71 1 2.94 3 8.83 1 2.94 Medium 4 11.76 1 2.94 1 2.94 1 2.94 Small 1 2.94 0 0.00 0 0.00 0 0.00 Undet. 14 41.18 0 0.00 2 5.88 0 0.00 TOTAL 24 70.59 2 5.88 6 17.65 2 5.88
Flesh removal Evisceration Fur removal Disarticulation
Pagina 1
Tab. 6 - Butchering operations sub-divided for prey size. NR: number of remains.
Foglio1
Size NR with CM % CM % Tot
Big 10 29.41 0.87 Medium 6 17.65 0.52 Small 1 2.94 0.09 Undetermined 17 50.00 1.49 TOTAL 34 100.00 2.97 Pagina 1
Tab. 5 - Abundance of cut-marks subdivided for prey size.
Fig. 3 - Cut-marks; a: location of cut-marks on Ursus spelaeus; b: fossil vertebra with cut mark; c: fossil rib with cut mark.
Foglio1 Taphonomic process NR % Deposition Mn oxyde 1142 99.83 Roots alteration 730 63.81 Carnivores intervention 627 54.81 Cracking 578 50.52 Trampling 365 31.9
Hydric abrasion - rounding 326 28.49 Hydric abrasion - smoothing 326 28.49 Limestone concretion 259 22.46
Cut marks 34 2.97
Pagina 1
Tab. 4 - Percentage of taphonomic processes. NR: number of re-mains.
morphic rock derived from quartz. The metamor-phic process causes recrystallization and most or all of the original texture and sedimentary structures of the stone are erased (Arzarello et al. 2011). The analysis conducted also confirms that the cut-marks obtained through simple quartz flakes during the experimental activities present the same characteris-tics of those observed in the archaeological record of the Ciota Ciara cave.
The Ciota Ciara cave deposit has undergone many taphonomic modifications. The deposit of manganese oxide on the whole surface of the re-mains and with deep penetration on the bones sug-gests a local environment characterized by caver-nous conditions of darkness and high humidity. In this environment, the abundant presence of super-ficial roots alterations can be attributable to the in-fluence of outside environment (i.e. roots of plants that grew out of the cave) (Morlan 1980; Grayson 1988). Water action was important for the forma-tion of the Ciota Ciara archaeological deposit; the presence of different degrees of hydraulic altera-tions, with combination of rounded surfaces and
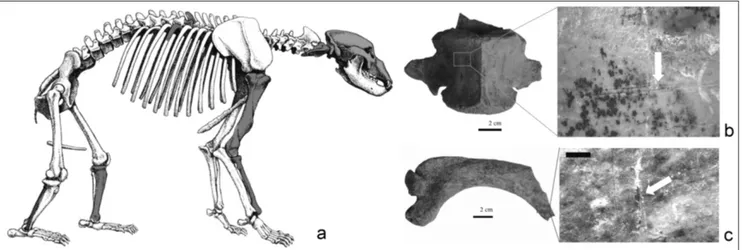
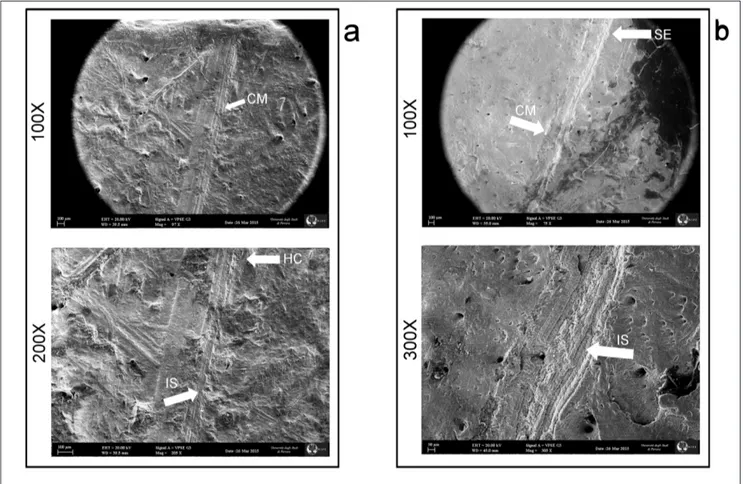
polish, identifiable on the remains allows us to assu-me the influence of a persistent transport with low energy, as confirmed by the lack of orientation of the finds. In the sample there are few finds showing a more marked degree of mineralization, and also that they were probably transported into the atrium from the inside of the cave by water. The concre-tions are due to precipitation of carbonate from the percolating water (Angelucci et al. 2015); in some cases, the concretions are subsequent the manga-nese deposit. The trampling marks observed on the Ciota Ciara fossil assemblage are characterized by multiple striae randomly distributed on the bone Fig. 4 - SEM images of cut-marks on fossil specimens from the Ciota Ciara Cave; a: cut mark on cave bear skull; b: cut mark on cave bear
vertebra. CM: cut-marks; IS: internal streaking; HC: herzian cones; SE: shoulder effect.
Foglio1
Characteristics Flint Quartz Quartzite
Slope of the edge abrupted half-abrupted variable Bottom section V-shaped V-shaped U-shaped Internal streaking thin wide wide Trait single multiple irregular Termination Y- shaped Y- shaped \
Shoulder effect X X X
Herzian cones X \ \
Pagina 1
surface. The intense trampling activity is probably due to the frequent occupation of the cave by U. spelaeus (Asryan et al. 2014), as attested by presence
of numerous cubs (Berto et al. 2016).The presen-ce of carnivore interventions with bite on fracture edge and the absence of epiphysis could suggest both primary and secondary intervention by carni-vores, with intense carcasses consumption.
The position and type of cut-marks are suf-ficient to assign the butchery activities. The data emerging from the archaeozoological studies prove that in the Ciota Ciara cave U. spelaeus was
butche-red by the Neanderthals and four activities can be observed: extraction of flesh masses, fur removal, disarticulation and evisceration. Concerning U. spe-laeus, activity of fur removal was attested (Auguste
1995; Munzel & Conard 2004; Armand 2006) by the presence of cuts-marks on a skull and a scapholu-nar of U. spelaeus. U. spelaeus faunal remains present
also traces of flesh extraction (Munzel & Conard 2004; Nilssen 2000). Those activities are attested by different traces: one scraping trace on the ven-tral side of a thoracic vertebra (vertebral body) and various scarification traces on two humeri and two tibias placed near the insertions of the muscle mas-ses. Finally, the presence of cut-marks on the atlas dorsal face can be related to disarticulation or fille-ting activities (Nilssen 2000).
Concerning C. lupus an activity of
disarticula-tion or filleting was attested by the presence of cut-marks on the third cervical vertebra (Nilssen 2000).
The defleshing of U. spelaeus by the
Neander-thals is attested in few other European Middle Pa-leolithic sites: Biache-saint-Vaast (France) (Auguste 1995), Taubach (Germany) (Bratlund 1999), Scladi-na cave in Belgium (Abrams et al. 2014) and Ni-etoperzowa Cave in Poland (Wojtal 2007). In Scladi-na cave, in layer 5, cut-marks on different fragments of long bones were identified, but they were associ-ated at the “chaîne opératoire” linked to bone working
(Abrams et al. 2014). Mousterian sites with evidence of fur removal on U. spelaeus are Biache-Saint-Vaast
(France) (Auguste 1995), La Madonna dell’Arma (Italy) (Cauche 2007), Taurach (Germany) (Bratlund 1999), Nietoperzowa Cave (Poland) (Wojtal 2007).
The data emerging from the taphonomic and archaeozoological studies of the S.U. 14 of the Ci-ota Ciara cave are in agreement with previous stud-ies already published (Arzarello et al. 2012; Arza-rello et al. 2013; Arnaud et al. 2014; Angelucci et al.
2015; Daffara et al. 2014; Berto et al. 2016). S.U. 14 is characterized by a lithic industry made with local raw materials with some elements made with
alloch-Fig. 5 - Cut-marks made with different raw material at optical micro-scope; a: cut mark made with flint; b: cut mark made with vein quartz; c: cut mark made with quartzite.
thonous flint. The Neanderthal occupation in this S.U. has been defined as “more intense” than that of the S.U. above (Arnaud et al. 2014; Angelucci et al. 2015; Daffara et al. 2014). This interpretation is based on the presence of a hearth and on the greater differentiation of the raw materials.
c
onclusIonsThis preliminary analysis demonstrates that it is possible to recognize the cut-marks left on
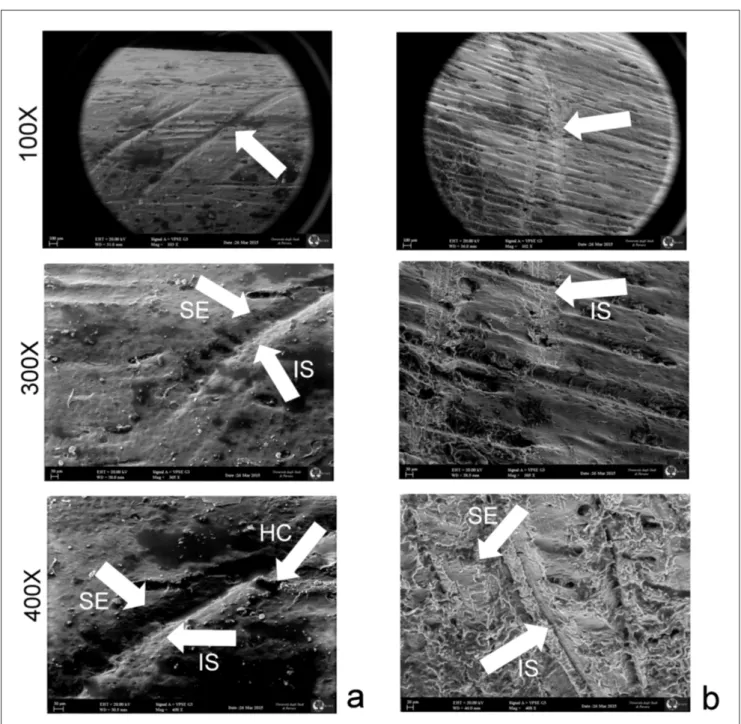
bones surfaces by simple flakes made of different raw materials. In order to make this study more de-tailed and complete, it will be necessary to increase the experimental database and to include different typologies of tools in future works. Nevertheless, the possibility of obtaining information about the raw material used for butchering activities from the analysis of the cut-marks could be a useful instru-ment for future investigations. This kind of analysis can be useful in areas where the use of flint was scarce or absent, like the Piedmont area (Daffara et al. 2014; Berruti et al. 2016). In sites where the use Fig. 6 - SEM images of cut-marks made with different raw material; a: cut mark made with vein quartz; b: cut mark made with quartzite. IS:
of quartz was limited compared to other raw mate-rials, a review of the cut-marks could reveal a more conspicuous use of quartz than previously sup-posed. The Ciota Ciara cave was intensely used as den by cave bear, as testified by the numerous cubs’ bone remains. The abundance of cave bear remains in all the layers suggests that this species used the cave for long periods. The presence of cut-marks on cave bear bones suggests that the chronologi-cal partition of the occupation of the cave between men and bears, as suggested in previous studies (i.e. Daffara et al. 2014), wasn’t that exclusive. The few bone remains with cut-marks suggest that bear ex-ploitation was not intensive. The cave bear exploi-tation by Neanderthals is attested in some others European sites, but the Ciota Ciara cave represents the first Italian record of this activity.
Acknowledgements. Our work has been possible thanks to the collaboration and support of the Borgosesia municipality, the GASB and the Soprintendenza Archeologia del Piemonte. Some material has been revised with the collaboration of Benedetto Sala who also hel-ped us on the interpretation of the faunal assemblage. Isabel Cáceres thanks Ministry of Economy and Competitiveness (MINECO) of Spain Government project nº CGL2015-65387-C3-1-P (MINECO/ FEDER) and was developed within the frame of the projects SGR 2014-899 (AGAUR, Generalitat de Catalunya), 2014PFR-URV-B2-17and 2015PFR-URV-B2-17 (Universitat Rovira i Virgili). Gabriele Berruti thanks the International Doctorate in Quaternary and Prehi-story (IDQP)’s scholarship. We also thank Margherita Zona Sara Daffara, Amerigo Barzaghi, Fabio Bertone and the three anonymous reviewers for their help.
RefeRences
Abrams G., Bello S.M., Di Modica K., Pirson S. & Bonjean D. (2014) - When Neanderthals used cave bear (Ursus spe-laeus) remains: Bone retouchers from unit 5 of Scladina Cave (Belgium). Quatern. Intern., 326-327: 274-287. Andrews P. (1990) - Owl, caves and Fossils. University of
Chi-cago Press,London, 239 pp.
Angelucci D.E., Arnaud J., Arzarello M., Berruti G.L.F., Berruto G., Berté D., Berto C., Buccheri F., Casini A.I., Daffara S., Luzi E., López-García J.M., Peretto C. & Zambaldi M. (2015) - Borgosesia, Monte Fenera. L’occupazione musteriana della grotta della Ciota Ciara, nuovi dati dalla campagna di scavo 2014. Quaderni della Soprintendenza Archeologica del Piemonte, 30: 400-402. Armand D. (2006) - Abri Castanet (Dordogne, France): An
aurignacian site with bear procurement. Bear explota-tion in Paleolithic time. Scientific Annals, School of Geology Aristotle University of Thessaloniki, 98: 263-268.
Arnaud J., Arzarello M., Berruti G., Berruto G., Bertè D., Berto C., Buccheri F., Casini A., Daffara S., Luzi E.,
Lopez Garcia J.M. & Peretto C. (2014) - Borgosesia, Monte Fenera. Grotta della Ciota Ciara. Nuovi dati sull’occupazione musteriana. Quaderni della Soprintendenza Archeologica del Piemonte, 29: 148-150.
Arzarello M., Marcolini F., Pavia G., Pavia M., Petronio C., Petrucci M., Rook L. & Sardella R. (2007) - Evidence of earliest human occurrence in Europe: the site of Pirro Nord (Southern Italy). Naturwissenschaften, 94(2): 107-112.
Arzarello M., Fontana F. & Peresani M. (2011) - Manuale di tecnologia litica preistorica. Carocci editore, Roma, 263 pp.
Arzarello M., Daffara S., Berruti G., Berruto G., Berté D., Ber-to C., Gambari F.M. & PeretBer-to C. (2012) - The Mouste-rian Settlement in the Ciota Ciara Cave: The Oldest Evi-dence of Homo neanderthalensis in Piedmont (Northern Italy). J. Biol. Res., 85: 71-75.
Arzarello M., Daffara S., Berruti G.L.F., Berruto G., Berté D.F., Berto C. & Peretto C. (2013) - Borgose-sia, Monte Fenera. Grotta della Ciota Ciara. Nuovi dati sull’occupazione musteriana e aspetti divulgativi. Quaderni della Soprintendenza Archeologica del Piemonte, 28: 300-302.
Asryan L., Ollé A. & Moloney N. (2014) - Reality and confu-sion in the recognition of post-depositional alterations and use-wear: an experimental approach on basalt tools. J. Lithic Studies, 1(1): 9-32.
Auguste P. (1995) - Chasse et charognage au Paléolithique moyen: l’apport du gisement de Biache-Saint-Vaast (Pas-de-Calais). Bull. Soc. Préhist. Franç., 92: 155-168.
Auguste P. (2002) - Fiche éclats diaphysaires du Paléolithique moyen: Biache-Saint-Vaast (Pas-de-Calais) et Kulna (Moravie, République tchèque). In: Patou-Mathis M. (Ed.) - Retouchoirs, compresseurs, percuteurs. Os à im-pressions et éraillures, Société Préhistorique Française, Fiches de la Commission de nomenclature sur l’industrie de l’os préhistorique, Cahier X: 39-57.
Bello S. M. & Soligo C. (2008) - A new method for the quanti-tative analysis of cutmark micromorphology. J. Archaeol. Sci., 35: 1542-1552.
Bello S.M., Parfitt S.A. & Stringer C. (2009) - Quantitative mi-cromorphological analyses of cut-marks produced by ancient and modern handaxes. J. Archaeol. Sci., 36: 1869-1880.
Bello S.M., Verveniotou E., Cornish L. & Parfitt S.A. (2011) - 3-dimensional microscope analysis of bone and tooth surface modifications: comparisons of fossil specimens and replicas. Scanning, 33(5): 316-324.
Berruti G. & Arzarello M. (2012) - L’analisi tracceologica per la ricostruzione delle attività nella preistoria: l’esempio della Grotta della Ciota Ciara (Borgosesia, VC). Ann. Univ. Ferrara, Museol. Sci. Nat., 8: 117-124.
Berruti G.L.F., Bertè D.F., Caracausi S., Daffara S., Ferreira C., Garanzini F., Rubat-Borel F. & Scoz L. (2016) - New evi-dences of human frequentations in the Alps: the project “Survey Alta Valsessera (Piedmont – Italy)”. Quatern. Intern., 402: 15-25.
neander-taliani della grotta della Ciota Ciara (Borgosesia, VC): analisi della provenienza delle materie prime. Master thesis, University of Ferrara.
Berto C., Bertè D.F., Luzi E., López-Garcia J.M., Pereswiet-Soltan A. & Arzarello M. (2016) - Small and large mam-mals from Ciota Ciara cave (Borgosesia, Vercelli, Italy): an Isotopic Stage 5 assemblage. Comptes Rendus Palevol, 15: 669-680.
Binford L.R. (1981) - Bones. Ancient Men and Modern Myths. Academic Press, New York, 320 pp.
Boëda E. (1988) - Le concept laminaire: rupture et filiation avec le concept Levallois. L’Homme de Néanderthal, L’Homme de Néandertal, vol. 8: la mutation: 41-59, ERAUL 35, Liège.
Boëda E. (1993) - Le débitage discoïde et le débitage Levallois récurrent centripète. Bull. Soc. Préhist. Franç., 90: 392-404. Bratlund B. (1999) - Taubach revisited. Jahr. Römisch-German.
Zentralmus. Mainz, 46: 67-174.
Brugal J.-P. & Raposo L. (1999) - Foz do Enxarrique (Ródão, Portugal): Preliminary results of the analysis of a bone assemblage from a Middle Palaeolithic open site. Monogr. Römisch-German. Zentralmus. Mainz, 42: 367-379.
Buccheri F. (2014) - Analisi tafonomica dei resti paleontologici dell’US 14 della Grotta della “Ciota Ciara” (Paleolitico Medio), Borgosesia - VC - Italia. Thesis Master Eras-mus Mundus em Quaternario e Préhistoria, University of Ferrara.
Bunn H.T. (1986) - Patterns of skeletal representation and hominid subsistence activities at Olduvai Gorge, Tan-zania, and Koobi Fora, Kenya. J. Human Evol., 15(8): 673-690.
Bunn H.T., Harris J.W.K., Isaac G.L., Kaufulu Z., Kroll E.M., Schick K., Toth N. & Behrensmeyer A.K. (1980) - FxJj50: an early Pleistocene site in northern Kenya. World Archaeology, 12: 109-136.
Cáceres I., Bennàsar M., Huguet R., Rosell J., Saladié P., Al-lué E., Solé A., Blasco R., Campeny G., Esteban-Nadal M., Fernández-Laso C., Gabucio M. J., Ibáñez N., Mar-tín P., Muñoz L. & Rodríguez-Hidalgo A. (2012) - Ta-phonomy of Level J of Abric Romaní. In: Carbonell i Roura. E. (Ed.) - High Resolution Archaeology and Neanderthal Behavior. Time and Space in Level J of Abric romaní (Capellades, Spain): 159-185. Vertebrate Paleobioloby and Paleoanthropology Series. Springer Science+Business.
Cauche D. (2007) - The Mousterian cultures in italian Ligu-ria: Analysis of lithic material of three sites in cave. L’Anthropologie, 111: 254-289.
Choi K. & Driwantoro D. (2007) - Shell tool use by early mem-bers of Homo erectus in Sangiran, central Java, Indonesia: cut mark evidence. J. Archaeol. Sci., 34: 48-58.
Clemente I. & Gibaja J.F. (2009) - Formation of use-wear traces in non-flint rocks: the case of quartzite and rhyo-lite. Differences and similiraties. In: Sternke F., Eigeland L. & Costa L.-J. (Eds) - Non-flint Raw Material Use in Prehistory. Old prejudices and new directions. UISPP - Proceedings of the World Congress (Lisbon, 4-9 Sep-tember 2006), vol 11. B.A.R. International Series Vol.
1939: 93-98. Archaeopress, Oxford.
Colonge D. & Mourre V. (2009) - Quartzite et quartzites, as-pects pétrographiques, économiques et technologiques des matériaux majoritaires du Paléolithique ancien et moyen du Sud-Ouest de la France. In: Grimaldi S. & Cura S. (Eds) - Technological analysis on quartzite ex-ploitation - Analyse technologique de l’exex-ploitation du quartzite, session WS15, XVIème Congrès de l’UISPP -Lisbonne, septembre 2006. B.A.R International Series Vol. 1998: 3-12. Oxford.
Daffara S., Arzarello M., Berruti G., Berruto G., Bertè D. & Berto C. (2014) - The Mousterian lithic assemblage of the Ciota Ciara cave (Piedmont, Northen Italy): exploi-tation and conditioning of raw material. J. Lithic Studies, 1(2): 63-78.
de Juana S., Galán A.B. & Domínguez-Rodrigo M. (2010) - Taphonomic identification of cut-marks made with lithic handaxes: an experimental study. J. Archaeol. Sci., 37: 1841-1850.
de Lumley H. (1972) - La Grotte moustérienne de l’Hortus. Etud. Quatern., 1: 527-624.
d’Errico F. & Giacobini G. (1986) - Orso delle caverne e uomo di Neandertal: fortuna di un mito e ricerca stratigrafica di una realtà. In: d’Errico F. & Giacobini G. (Eds) - I cacciatori neandertaliani: 124-127. Jaca Book, Milano. Domínguez-Rodrigo M., de Juana A., Galán B. & Rodriguez
M. (2009) - A new protocol to differentiate trampling marks from butchery cut-marks. J. Archaeol. Sci., 36: 2643-2654.
Fernández-Jalvo Y. & Andrews P. (2003) - Experimental Ef-fects of Water Abrasion on Bone Fragments. J. Taphon., 1(3): 147-163.
Fernández-Jalvo Y. & Cáceres I. (2010) - Tafonomía e Indu-stria Lítica: marcas de corte y materias primas: 277-290 . Cuaternario y Arqueología: Homenaje a Francisco Giles Pacheco, Cádiz.
Forestier H. (1993) - Le Clactonien: Mise en application d’une nouvelle méthode de débitage s’inscrivant dans la vari-abilité des systèmes deproduction lithique du Paléo-lithique ancien. Paléo, 5: 53-82.
Gabori-Csank V. (1968) - La station du Palèolithique moyen d’Erd, Hongrie. Akademia Kiado, Budapest, 277 pp. Galán A.B. & Domínguez-Rodrigo M. (2013) - An
experimen-tal study of the anatomical distribution of cut-marks created by filleting and disarticulation on long bone ends. Archaeometry, 55(6): 1132-1149.
Giacobini G. (1995) - Identificazione delle tracce di macel-lazione con strumenti litici. Analisi di microscopia elettronica a scansione. Atti del 1 Convegno Nazionale di Archeozoologia. Rovigo - Accademia dei Concordi 5-7 Marzo 1993, 1: 29-37. Padusa Quaderni, Padova. Grayson D.K. (1988) - Danger Cave, Last Supper Cave, and
Hanging Rock Shelter: the Faunas. Ame. Mus. Nat. Hist. Anthropol. Pap., 66: 1-130.
Greenfield H.J. (1999) - The origin of metallurgy: distinguish-ing stone from metal cut-marks on bones from archaeo-logical sites. J. Archaeol. Sci., 26:707-808.
diagnostics for identifying stone tool type and raw mate-rial. J. Field Archaeol., 31: 147-163.
Igreja M. (2009) - Use-wear analysis of non-flint stone tools using DIC microscopy and resin casts: a simple and ef-fective technique. In: Recent functional studies on non flint stone tools: methodological improvements and ar-chaeological inferences. Proceedings of the workshop. [CD-ROM]. Lisboa (Padrão dos Descobrimentos), 2009.ISBN978-989- 20-1803-4/http://www.workshop-traceologia-lisboa2008.com/
Jéquier C.A., Romandini M. & Peresani M. (2012) - Les re-touchoirs en matières dures animales: une comparaison entre Moustérien final et Uluzzien. Comptes Rendus Pa-levol, 11: 283-292.
Koby F.E. (1951) - Grottes autrichiennes avec culte de l’ours? Bull. Soc. Préhist. Franç., 48: 8-9.
Kurtén B. (1958) - Life and death of the Pleistocene cave bear. Acta Zool. Fennica, 95: 1-56.
Lartet E. & Christy H. (1875) - Reliquiae Acquitancae: being contributions to the archaeology and paleontology of Perigord and adjoining provinces of Southern France. Williams and Norgate, London, 722 pp.
Lemorini C., Plummer T.W., Braun D.R., Crittenden A.N., Ditchfield P.W., Bishop L.C., Hertel F., Oliver J.S., Mar-lowe F.W., Schoeninger M.J. & Potts R. (2014) - Old stones’ song: Use-wear experiments and analysis of the Oldowan quartz and quartzite assemblage from Kanjera South (Kenya). J. Hum. Evol., 72: 10-25.
Lombera Hermida A. (2008) - Quartz morphostructural groups and their mechanical implications. Ann. Univer. Studi Ferrara, Museol. Sci. Natural., Vol. spec.: 101-105. López-Gonzalez F., Grandal-d’Anglade A. & Vidal-Romani
J.R. (2006) - Deciphering bone depositional sequences in caves through the study of manganese coatings. J. Ar-chaeol. Sci., 33: 707-717.
Marín-Arroyo A. B., Landete-Ruiz M. D., Seva-Román R. & Lewis M. D. (2014) - Manganese coating of the Tabun faunal assemblage: Implications for modern human be-haviour in the Levantine Middle Palaeolithic. Quatern. Intern., 330: 10-18.
Martin M.H. (1909) - Traces de Coups de Silex sur la Machoire Inférieure des Ruminants et des Équidés a la Période Moustérienne. Association Française pour l’Avancement des Sciences, Paris. (7 août): 777-782.
Martin M.H. (1910) - Traces humaines laissées sur les os a l’époque moustérienne. Constatations faites dans le gisement de la Quina (Charente). Association Française pour l’Avancement des Sciences, Toulouse (3 août): 242-245.
Moretti E., Arrighi S., Boschin F., Crezzini J., Aureli D. & Ronchitelli A. (2015) - Using 3D microscopy to analyze experimental cut-marks on animal bones produced with different stone tools. Ethnobiol. Lett., 6(2): 267-275. Morlan R.E. (1980) - Taphonomy and archaeology in the
Upper Pleistocene of the northern Yukon Territory: a glimpse of the peopling of the New World. National Museums of Canada Ottawa, 398 pp.
Mourre V. (1996) - Les industries en quartz au Paléolithique. Terminologie, méthodologie et technologie. Paléo, 8: 205-223.
Munzel S.C. & Conard N.J. (2004) - Cave bear hunting in Höh-le Fels Cave in the Ach ValHöh-ley of the Swabian Jura. Rev. Paléobiol., 23: 877-885.
Nilssen P.J. (2000) - An actualistic butchery study in South Africa and its implications for reconstructing hominid strategies of carcass acquisition and butchery in the Up-per Pleistocene and Plio-Pleistocene. PhD thesis, De-partament of Archaeology, University of Cape Town. Oliver J.S. (1989) - Analogues and site context:bone
damag-es from Shield Trap Cave (24 CB91), Carbon County, Montana, U.S.A. In: Bonnichsen R. & Sorg M.H. (Eds) - Bone modification: 73-98. Center for the Study of the First American, Orono.
Olsen S.L. & Shipman P. (1988) - Surface Modification on Bone: Trampling versus Butchery. J. Archaeol. Sci., 15: 535-53.
Palma di Cesnola A. (2006) - L’Aurignacien et le Gravet-tien Ancien de la Grotte Paglicci au Mont Gargano. L’Anthropologie, 110: 355-370.
Schick K. & Toth N. (1993) - Making silent stones speak. Si-mon & Schuster, New York, 351 pp.
Shipman P. (1983) - Early hominid lifestyle: hunting and gath-ering or foraging and cavenging? In: Clutton-Brock J. & Grigson C. (Eds) - Animals and archaeology, 1: hunters and their prey: 31-9. International Series, British Archae-ological Reports, Oxford.
Shipman P. (1986) - Scavenging or hunting in early hominids: theoretical framework and tests. American Anthropologist, 88: 27-43.
Shipman P. & Rose J. (1983) - Early Hominid Hunting, Butch-ering, and Carcass-Processing Behaviors: Approaches to the Fossil Record. J. Anthropol. Archaeol., 2: 57-98. Shipman P., Fisher D.C. & Rose J.J. (1984) - Mastodon
butch-ery: microscopy evidence of carcass processing and bone tool use. Paleobiology, 10: 358-365.
Valensi P. & Psathi E. (2004) - Faunal exploitation during the Middle Palaeolithic in south-eastern France and north-western Italy. Intern. J. Osteoarchaeol., 14: 256-272. Walker P.L. & Long J. C. (1977) - An experimental study of
the morphological characteristics of tool marks. Ameri-can Antiquity, 42, 605-616.
West J.A. & Louys J. (2007) - Differentiating bamboo from stone tool cut-marks in the zooarchaeological record, with a discussion on the use of bamboo knives. J. Ar-chaeol. Sci., 34: 512-518.
Wojtal P. (2007) - Zooarchaeological studies of the Late Pleis-tocene sites in Poland. Polish Academy of Sciences, 189 pp.
Yravedra J., Dominguez-Rodrigo M., Santonja M., Pérez-González A., Panera J., Rubio-Jara S. & Baquedano E. (2010) - Cut-marks on the Middle Pleistocene elephant carcass of Áridos 2 (Madrid, Spain). J. Archaeol. Sci., 37: 2469-2476.