UNIVERSITÁ DEGLI STUDI DI CATANIA
FACOLTÁ DI SCIENZE MATEMATICHE FISICHE E NATURALI
SEZ. DI BIOLOGIA ED ECOLOGIA VEGETALE (D.A.C.P.A)
DOTTORATO DI RICERCA IN BIOLOGIA ED ECOLOGIA VEGETALE IN
AMBIENTE MEDITERRANEO – XXIII CICLO
SEDE
AMMINISTRATIVA:
UNIVERSITÁ
DEGLI
STUDI
DI
CATANIA
Daniela Cataldo
I licheni epifiti dei SIC “Bosco del Flascio” e “Rocche di Roccella
Valdemone”, (Sicilia, nord orientale): interpretazione ecologica e
chiavi analitiche.
TESI DI DOTTORATO
TUTOR: COORDINATORE:
DOTT. SSA MARIA GRILLO PROF. GIUSEPPE MAUGERISommario………pag.
1Riassunto……….…...4
2 Introduzione...5
2.1 Aree protette e salvaguardia dei boschi………...………...6
2.2 I licheni epifiti dei SIC Bosco del Flascio e Rocche di Roccella Valdemone: stato dell’arte…...………5
2.3 Il progetto………..………6
3 Le aree di studio………..………...7
3.1 I Monti Nebrodi………..8
3.2 I SIC “Bosco del Flascio” e “Rocche di Roccella Valdemone”: caratteristiche stazionali………...10
4 Impatto antropico nella regione mediterranea………...………...13
4.1 Impatto antropico nei SIC………..………..15
5 Il clima e il bioclima……….………17
6 I licheni………..…….……….19
6.1 Storia dell’uso dei licheni………...21
6.2 Il declino delle comunit{ licheniche………...23
7 Materiali e metodi……….……….…...25
7.1 Metodo di campionamento……….……….25
8 La flora lichenica dei SIC………...…………...27
8.1 Elenco floristico dei licheni epifiti del SIC Bosco del Flascio………..27
8.2 Elenco floristico dei licheni epifiti del SIC Rocche di Roccella Valdemone...30
8.3 Commento alla flora……….………...33
8.4 Distribuzione delle specie in base ai piani fitoclimatici……….37
9 Il concetto di autoecologia………..……….………38
9.1 Autoecologia dei licheni.………...………40
9.2 I fotobionti, eccetto i cianobatteri………...41
9.3 I cianobatteri………..………..42
9.4 I cianolicheni del SIC Bosco del Flascio………...44
9.5 I cianolicheni del SIC Rocche di Roccella Valdemone……….46
12 I macrolicheni………..……….………56
13 Biomonitoraggio……….………..……….……….………59
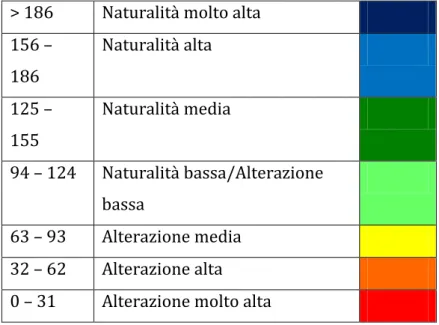
13.1 L’Indice di Biodiversit{ Lichenica (I.B.L.)……….……….………. 60
13.2 La Biodiversità Lichenica, BL, nei SIC………...61
13.3 Commento ai dati di BL ottenuti……….……...63
13.4 PLOT SIC BOSCO DEL FLASCIO...……….………..64
13.5 PLOT SIC ROCCHE DI ROCCELLA VALDEMONE………..………71
14 Considerazioni conclusive………...76
Bibliografia………...……….80
Appendice…………..………....88
- I licheni epilitici e di altri substrati del SIC Bosco del Flascio………...88
- I licheni epilitici e di altri substrati del SIC Rocche di Roccella Valdemone...89
- Chiave analitica dei licheni del SIC Bosco del Flascio………...90
- Chiave analitica dei licheni del SIC Rocche di Roccella Valdemone………....96
- Tavole iconografiche………..102
- Allegati………..….146 Ringraziamenti
1 Riassunto
Obiettivo di questa ricerca è stato quello di ricavare quante più informazioni possibili dal censimento della flora di due SIC ubicati sui Monti Nebrodi, i quali sono molto simili per caratteristiche stazionali, ma piuttosto diversi per alcune caratteristiche ambientali e per la gestione.
Si è voluto valutare se le informazioni ricavabili dall’autoecologia dei licheni fossero indicazioni indirette dell’ecologia dei siti considerati. La risposta è stata positiva, la ricerca ha mostrato diverse congruenze tra le informazioni ricavate e quelle disponibili. In particolare: presenza di disturbo antropico, microclima umido e gestione sono risultati essere i fattori più importanti nella selezione di una flora epifita qualitativamente ricca di specie, ma soprattutto indicatrice di continuità ecologica.
Abstract
The aim of this research was to obtain as much information as possible from the census of the flora of two SIC located on the Mounts Nebrodi, which are very similar in features stational, but rather different for certain environmental characteristics and management. The response was positive, the research showed several matches between the information obtained and available. In particular: the presence of human disturbance, humid microclimate and management were found to be the most important factors in selecting a quality epiphytic flora rich in species, but also an indicator of ecological continuity.
2 Introduzione
2.1 Aree protette e salvaguardia dei boschi.
Negli ultimi decenni sono stati condotti diversi studi riguardanti la “salute degli ecosistemi forestali” i quali hanno messo in evidenza l’esistenza di preoccupanti modificazioni ambientali causate in primo luogo dai cambiamenti d’uso del territorio e conseguentemente da tutte le attività connesse. In modo particolare i boschi sono stati salvaguardati per mezzo di piani di ricerca approfonditi (ICP IM “International Cooperative Programme on Integrated Monitoring of Air Pollution Effects on Ecosystems” e CON.ECO.FOR. “Controllo Ecosistemi Forestali”) e dell’istituzione di aree protette.
Salvaguardare i boschi e le aree di grande valore naturalistico diventa anche sul territorio italiano un obiettivo da raggiungere: la prima area protetta fu istituita il 3 dicembre 1922; era il Parco Nazionale del Gran Paradiso ubicato tra il Piemonte e la Valle d’Aosta il primo parco nazionale italiano.
La prima area protetta ad essere istituita in Sicilia fu il Parco dell’Etna, il 17 marzo del 1987. Successivamente, in accordo con quanto stabilito dalla Commissione Europea in materia di protezione dell’ambiente con le direttive: Direttiva n. 79/409/CEE Uccelli e Direttiva 92/43/CEE Habitat, sono stati istituiti Siti di Importanza Comunitaria (SIC), Zone di Protezione Speciale (ZPS), Aree Marine Protette (AMP), Importants Birds Areas (IBA) e negli ultimi tempi Important Plants Areas (IPAs).
Nel giugno del 1995 il Ministero dell'Ambiente e della Tutela del Territorio e del Mare attuò formalmente il progetto “Bioitaly” con l’obiettivo di recepire e dare concreta attuazione alle Direttive Europee "Habitat" ed "Uccelli”. Tale progetto ha previsto la raccolta e la sistematizzazione delle informazioni sui biotopi, sugli habitat naturali e seminaturali di interesse comunitario e sulla loro collocazione geografica. Una volta identificati gli habitat, la loro collocazione e le specie di interesse comunitario, si è proceduto a segnalare tali aree, denominate Siti di Interesse Comunitario (SIC), alla Commissione Europea al fine della loro inclusione nella Rete Natura 2000.
In Sicilia, con decreto n. 46/GAB del 21 febbraio 2005 dell'Assessorato Regionale per il Territorio e l'Ambiente, sono stati istituiti 204 SIC, 15 ZPS, 14 aree contestualmente SIC e ZPS per un totale di 233 aree da tutelare.
2.2 I licheni epifiti dei SIC Bosco del Flascio e Rocche di Roccella Valdemone: stato dell’arte.
Numerosi studi riguardano i licheni di Sicilia, a partire da “Lichenographia sicula” di Tornabene del 1848 passando per gli studi riguardanti le flore, gli usi nella farmacopea, gli usi nella cosmesi sino a oggi con l’uso dei licheni come bioindicatori e bioaccumulatori (Ottonello et al., 2000; Basile et. al., 2008).
In argomento, è poi possibile consultare on line un database lichenologico (http://dbiodbs.univ.trieste.it/italic/italic02a), da cui in pochi step si possono conoscere le specie licheniche sinora censite in Italia1; a questo database si affianca il sito web
“Progetto KeytoNature – Dryades”, branca italiana del Progetto Europeo KeyToNature, dove è possibile visionare le flore licheniche di varie regioni italiane e non, e di utilizzare le relative chiavi analitiche comprese quelle dei due SIC esaminati, (http://www.dryades.eu/home1.html; Nimis et al., 2008).
Per quanto attiene alle notizie bibliografiche relative ai due SIC, poiché ubicati sui Monti Nebrodi, è possibile conoscerne da lavori relativi al territorio nebrodense; ad esempio si hanno notizie relative alla presenza di Lobaria pulmonaria nel Bosco del Flascio e nel Bosco di Malabotta poiché indagati nell’ambito del “Progetto Lobaria” su territorio nazionale (Nascimbene et al., 2006; 2008).
Si parla ancora di licheni dei Nebrodi in due articoli di Grillo (1992 e 1996) che annota per i territori di Capizzi e del Bosco di Malabotta rispettivamente 94 e 68 specie epifite.
Con questo lavoro si aggiornano le conoscenze dei licheni dell’area nebroidea e si apporta ai due SIC un nuovo contributo scientifico importante sia per il dato strettamente relativo alle flore che per le conoscenze apprese sull’influsso dei fattori ecologici sulla presenza e sulla qualità dei licheni che sono stati ritrovati.
2.3 Il progetto
Si è scelto di studiare le cenosi boschive di due aree protette: una denominata “Bosco del Flascio” ricadente nella provincia di Catania e l’altra “Rocche di Roccella Valdemone ”, in quella di Messina.
Queste due aree sono state scelte per la presenza di formazioni boschive a caducifoglie piuttosto estese, che si prestano all’obiettivo della nostra ricerca: descrivere attraverso la flora lichenica il grado di qualità ambientale e la continuità ecologica delle due cenosi.
Considerando il significato autoecologico dei licheni rinvenuti abbiamo elaborato un piano di studio che mette in risalto come il mantenimento di condizioni ecologiche piuttosto stabili influenzi positivamente la persistenza di specie vulnerabili o sensibili ai cambiamenti ambientali.
3 Le aree di studio
Le indagini lichenologiche sono state espletate su due soprassuoli boschivi che per caratteristiche stazionali e vegetazionali sono molto simili; complessivamente il territorio scelto si estende per circa 3.800 ha.
Entrambi i SIC sono ubicati sul versante meridionale dei Monti Nebrodi, nella Sicilia Nord-Orientale (Fig.1) e approssimativamente sono distanti l’uno dall’altro 13 km.
Gran parte del territorio del SIC “Bosco del Flascio” è demanio forestale, in gestione quindi all’Azienda Foreste Demaniali della Regione Sicilia; l’istituzione del SIC ha solamente inglobato quella che già era un’area sottoposta a vincoli; il SIC “Rocche di Roccella Valdemone” non ha a oggi un Piano di Gestione, nella stessa situazione si trova il SIC Bosco del Flascio. In queste aree sono di particolare interesse naturalistico e paesaggistico boschi mesofili a querce caducifoglie che ancora si mantengono in buono stato di conservazione - in misura di gran lunga superiore nel SIC Bosco del Flascio - e all’interno dei quali possono essere rinvenute delle specie vegetali di interesse biogeografico rare nell’area regionale, cui si accompagna una componente faunistica piuttosto interessante.
3.1 I Monti Nebrodi
I Monti Nebrodi si estendono per circa 70 km tra i Peloritani ad est e le Madonie ad ovest costituiscono l’Appennino Siculo.
Geologicamente (Fig. 2) questo territorio è costituito da varie successioni sedimentarie tra loro sovrapposte in cui è possibile riconoscere due successioni primarie, la numidica e la sicilide, le quali costituiscono rispettivamente tre e tre unità tettoniche principali: Maragone, Monte Salici- Monte Castelli, Nicosia e Monte Soro, Argille Scagliose superiori, Troina, (Duee, 1965). E’ presumibile che l’orogenesi di tipo compressivo dell’area nebroidea si sia protratta fino al Pliocene superiore, circa 2,5 milioni di anni fa (Bianchi et al. 1987) e abbia generato la sequenza di rilievi superanti tutti quasi tutti i 1.500 m di altezza e dalle cui vette si dipartono valli e crinali, la cui morfologia (Fig. 3) è nota a tutti.
Per quanto riguarda l’idrografia del versante meridionale dei Monti Nebrodi essa comprende una serie di corsi d’acqua quasi tutti a carattere torrentizio (Fig. 4) che durante il periodo delle piogge trasportano a valle grandi quantità di detriti; tra i corsi d’acqua di una certa importanza è il fiume Flascio che è il maggiore affluente di destra del fiume Alcantara e fa parte del reticolo idrografico superficiale del corpo idrico sotterraneo Monte Soro.
Fig. 5: Mappa del SIC Bosco del Flascio.
3.2 I SIC “Bosco del Flascio” e “Rocche di Roccella Valdemone”: caratteristiche stazionali.
I SIC ITA070007 “Bosco del Flascio” e ITA030034 “Rocche di Roccella “Valdemone” (Fig. 5 e 8) sono ubicati sul versante meridionale dei Monti Nebrodi. Il primo si estende su di un’area di 2948 ha, compresa per intero all’interno del Parco degli omonimi monti, le cui quote variano fra 900 e 1600 m e che coincide con l’alto bacino del fiume Flascio (Fig. 7).
Il territorio del SIC salvo le aree più scoscese è attraversabile grazie ai sentieri dell’ Azienda Foreste Demaniali, alla stessa appartengono diverse strutture rurali di riparo per le attrezzature e all’interno delle quali, previa autorizzazione, è possibile pernottare.
Il secondo si estende su di un territorio di 865 ha (Fig: 8) le cui quote sono comprese tra gli 800 e 1300 m. Il territorio del SIC si snoda tra ripidi pendii e affioramenti rocciosi.
Come si osserva dalle foto l’area (Figg. 9-10) boscata è frammentata, una parte si trova alla sommità del SIC dove è diffuso il castagno; nelle aree di S-O e S-E troviamo i querceti frammisti sempre al castano ed altre essenze, come i noccioleti.
Fig. 10: Paesaggio del versante di S-E. Fig. 9: Veduta panoramica del SIC.
4 Impatto antropico nella regione mediterranea.
L’impoverimento e la successiva scomparsa degli habitat a causa dell’impatto delle attività antropiche ha radici molto antiche: già durante il Medioevo, in Europa, si verificò un forte incremento demografico che comportò richieste sempre maggiori di risorse e al contempo ne causò un’anomala distribuzione; questo, tra i tanti accadimenti, portò anche al progressivo disboscamento delle aree forestali originarie a favore delle aree coltivate e provocò un massiccio sfruttamento del legname per le costruzioni.
Tuttavia sempre nel Medioevo, con l’avvento del monachesimo i boschi vennero implicitamente salvaguardati poiché luoghi che per l’immaginario collettivo erano popolati da mostri e fiere, quindi legati ad ancestrali paure ed di conseguenza evitati. Paradossalmente proprio in questo periodo nascono i primi “parchi naturali” legati alle attività di caccia riservate ai signorotti e precluse ai servi (Fig. 11); per questi luoghi riservati a pochi soggetti l’impatto antropico sull’ecosistema boschivo era, di fatto, molto contenuto. In seguito nacquero i primi giardini protetti urbani in contrapposizione agli inospitali boschi extraurbani.
La regione Mediterranea in particolare, per alcuni millenni è stata sottoposta a modifiche profonde, dovute principalmente alla distruzione delle foreste primitive che nel tempo sono state sostituite dalle comunità arbustive di macchia e gariga, (Pignatti in Holzner et al., 1983). Si presume che le attività umane nel bacino Mediterraneo siano iniziate nel periodo Neolitico con la domesticazione di piante e animali; 10.000 anni fa, al tempo dei Romani, si comincia a parlare di uso del territorio e dopo la caduta dell’Impero Romano e fino al 19esimo secolo, in particolare in Italia, si assiste ad una generale scomparsa delle aree boschive a favore dell’agricoltura e della pastorizia, attività questa che divenne l’occupazione più diffusa sul territorio; non meno importante da considerare fu il
(2004), oggi la presenza di aree naturali e seminaturali è piuttosto compromessa; esse interessano il territorio regionale per il 31,3% della superficie totale, in particolare i complessi boschivi veri e propri ne occupano solamente il 24% (ARTA, 2005).
La maggior parte del patrimonio naturalistico, con maggiore riguardo per i boschi, viene salvaguardato all’interno di aree protette regionali e in più di 70 habitat di interesse comunitario, sui 200 inclusi nell’allegato I della Dir. Habitat sono compresi nei Siti d’Importanza Comunitaria censiti per la Sicilia.
“Noi deploriamo oggi la frequenza delle inondazioni; e, se potessimo volgere a beneficio del rimboschimento delle nostre montagne il denaro che ogni anno si disperde per riparare ai dilagamenti dei nostri torrenti e fiumi, i gravi danni che dobbiamo lamentare sarebbero assai ridotti di numero e di quantit{” (Tip. Monauni, 1901).
“Basta per un momento solo riflettere che le specie silvane hanno assoluto bisogno dell’ombra, del verde e dei frutti delle foreste, per comprendere quale strazio nel mondo faunistico compia l’uomo disboscando. Se il cinghiale, il capriolo, il daino, il muflone, il cervo e tutti gli altri mammiferi e l’infinito stuolo di uccelli della foresta sono così diminuiti non solo in Italia, ma anche nel resto del mondo, dobbiamo attribuire gran parte della colpa proprio al taglio dei boschi” (Vaccari, 1912).
4.1 Impatto antropico nei SIC.
I Monti Nebrodi offrono ancora oggi al visitatore e allo studioso una Sicilia verde, ricca di boschi e oasi naturali da togliere il fiato, così il SIC Bosco del Flascio che tra i suoi sentieri mostra scorci incanti.
Tuttavia nel corso del tempo si è verificata una generale riduzione delle aree boscate nebrodensi, cosa che in parte può essere imputata alle coltivazioni agricole che, siano esse frutteti, campi di grano o uliveti, vengono impiantate sino anche i 1000 m di altitudine e insieme agli estesi pascoli, tipici di queste montagne, vanno a “circondare” le formazioni boschive.
Si tratta di formazioni a dominanza di Quercus cerris miste ad altre essenze arboree come Quercus virgiliana alle quote comprese tra gli 800 e i 1200 m, Acer
campestre e Fagus sylvatica alle quote più elevate; nel sottobosco si trovano arbusti di Prunus spinosa, Rosa canina, Crataegus monogyna…; lo strato erbaceo comprende specie
sia nemorali come Cyclamen hederifolium e Primula vulgaris che provenienti dai pascoli come Dactylis glomerata e Silene italica.
In una situazione di questo genere l’ingresso nei boschi di specie erbacee dai pascoli, dalle radure e dalle colture, testimonia quanto l’impatto antropico possa alterarne il corteggio floristico (Poli & Lo Giudice, 1988).
- Nel caso del SIC “Bosco del Flascio” il cui organo responsabile della gestione è per gran parte, l’Azienda Foreste Demaniali, le pratiche forestali per la tutela del bosco e l’assenza di animali al pascolo nelle aree di maggiore tutela (zona A-B), favoriscono la persistenza di condizioni di naturalità; interferisce con la qualit{ generale dell’habitat il passaggio di mezzi non autorizzati (quad, moto cross) sulle tracce carrabili e lungo i sentieri. A causa di questo passaggio si è determinata erosione del suolo, dovuta ai solchi lasciati dalle ruote che sono poi approfonditi dallo scorrimento delle acque meteoriche, e frastuono causato dai motori che disturba la fauna selvatica.
Ancora, diversi anni fa sono state costruite delle briglie in cemento lungo il corso del fiume e un serbatoio idrico a scopo irriguo; queste opere hanno ridotto sensibilmente la portata del corso d’acqua e hanno “imbruttito” il paesaggio; si potrebbero proporre di riqualificare il corso del fiume attraverso opportune opere di riqualificazione ambientale.
frequentemente utilizzato a pascolo da ovini e bovini e non di rado è percorso dal fuoco. Gli incendi sono una pratica “usuale” nella pastorizia per favorire la crescita di piante pabulari, non solo, in alcuni casi vengono appiccati come ammonimento in caso di conflitto con le guardie forestali o tra gli stessi pastori. A causa della ripetizione di questi eventi si è prodotto nel tempo sovra sfruttamento e erosione del suolo.
Dagli annali degli incendi del Corpo Forestale di Floresta (Messina) si apprende che negli ultimi cinque anni si sono verificati numerosi incendi che hanno interessato le aree più esterne del SIC e come si vede dalle carte allegate in appendice (Fig. 12 e 13) gli incendi sono stati appiccati da bordo strada e hanno risalito i versanti.
5 Il clima e il bioclima.
La Sicilia che è la più grande isola del Mediterraneo si estende per 25.000 kmq in latitudine tra i 36° e i 38° nord e in longitudine fra il 12° e il 15° est.
Nonostante il territorio sia geomorfologicamente piuttosto complesso è possibile suddividerlo in tre differenti versanti: quello settentrionale da Capo Peloro a Capo Lilibeo, quello meridionale da Capo Lilibeo a Capo Passero ed infine quello orientale da Capo Passero a Capo Peloro.
La complessità orografica che alterna porzioni montuose a collinari e di pianura, ne condiziona l’andamento climatico.
Se secondo la classificazione macroclimatica di Koppen (1936) la Sicilia può essere definita una regione a clima mediterraneo caratterizzato da una temperatura media del mese più caldo superiore ai 22 C° e da un regime delle precipitazioni concentrate nel periodo freddo (autunno-inverno), nel particolare la complessità morfologica del territorio complica le cose. Nelle diverse aree del territorio siciliano è possibile, infatti, distinguere diversi sottotipi climatici: clima temperato subtropicale, temperato caldo, temperato sublitoraneo, temperato subcontinentale, temperato fresco. Queste differenze non solo si riscontrano tra zone molto distanti e ad altitudini diverse, ma anche tra località a latitudine e altitudine piuttosto simili ma poste su versanti differenti.
L’analisi del clima per i due SIC è stata fatta sulla base dei dati pluviometrici tratti da Duro et al. (1996), mentre i dati relativi alle temperature, mancando nei siti le stazioni termometriche, sono stati prodotti col metodo SPSS (interpolazione lineare), che tiene conto dell’altezza sul livello del mare.
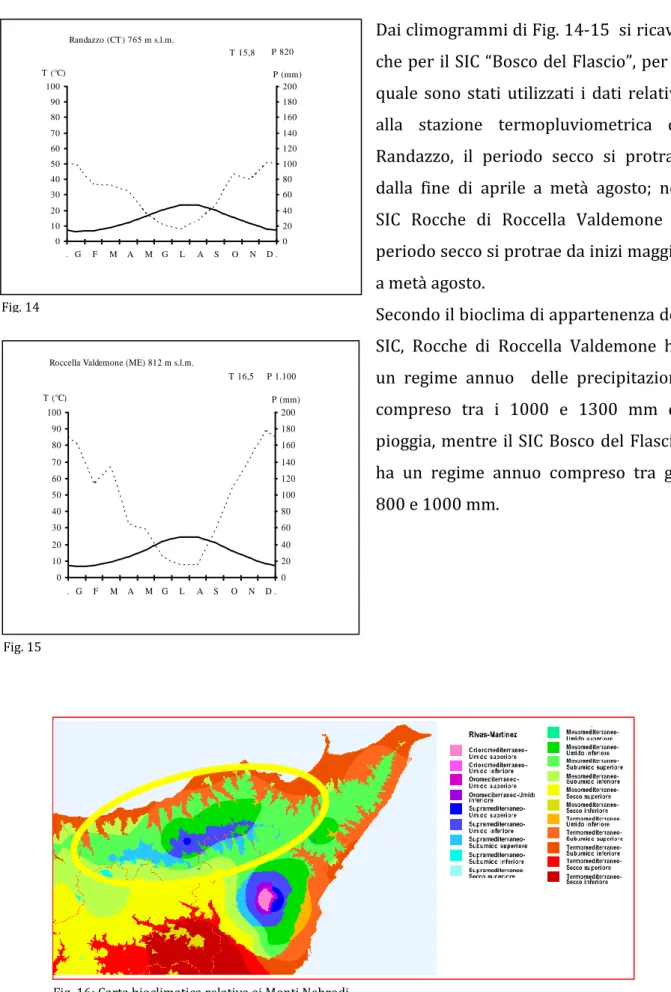
Questi dati vengono elaborati in un climogramma (Fig. 14-15) secondo Bagnouls & Gaussen, (1957) che viene costruito in sistema di assi cartesiani nel quale in ascissa sono riportati i mesi dell’anno e in ordinata a sinistra i valori medi della temperatura e a destra le precipitazioni riscontrate, secondo la formula T=2P.
L’intersezione della curva delle precipitazioni con quella delle temperature medie corrisponde al periodo arido. Il bioclima (Fig. 16), secondo la classificazione di Rivas Martinez (1982), del SIC “Bosco del Flascio” rientra nel tipo mesomediterraneo con ombrotipo subumido superiore; per quanto riguarda il SIC “Rocche di Roccella
Dai climogrammi di Fig. 14-15 si ricava che per il SIC “Bosco del Flascio”, per il quale sono stati utilizzati i dati relativi alla stazione termopluviometrica di Randazzo, il periodo secco si protrae dalla fine di aprile a metà agosto; nel SIC Rocche di Roccella Valdemone il periodo secco si protrae da inizi maggio a metà agosto.
Secondo il bioclima di appartenenza dei SIC, Rocche di Roccella Valdemone ha un regime annuo delle precipitazioni compreso tra i 1000 e 1300 mm di pioggia, mentre il SIC Bosco del Flascio ha un regime annuo compreso tra gli 800 e 1000 mm.
Fig. 16: Carta bioclimatica relativa ai Monti Nebrodi. Roccella Valdemone (ME) 812 m s.l.m.
0 10 20 30 40 50 60 70 80 90 100 . G F M A M G L A S O N D . P 1.100 T (°C) 0 20 40 60 80 100 120 140 160 180 200 T 16,5 P (mm) Fig. 15 Randazzo (CT ) 765 m s.l.m. 0 10 20 30 40 50 60 70 80 90 100 . G F M A M G L A S O N D . P 820 T (°C) 0 20 40 60 80 100 120 140 160 180 200 T 15,8 P (mm) Fig. 14
6 I licheni.
Il termine “licheni” ha origine greca e per la prima volta venne utilizzato nel IV secolo da Teofrastro, filosofo e botanico, il quale li indicava come “piante che crescono sui tronchi d’albero”, confondendoli probabilmente con delle epatiche. Da allora in poi viene fatto cenno sui licheni solo sporadicamente; nel 1867 finalmente Schwendener ne scopre la vera natura.
Dal punto di vista biologico un lichene è un “consorzio”, cioè una convivenza stabile e stretta tra specie diverse, un fungo e un’alga, nel particolare un individuo che è il micelio, costituito da un intreccio di ife, di un fungo e una numerosa colonia di alghe unicellulari, cioè numerosi individui di alga (gonidi).
Il nuovo organismo che si costituisce a seguito della simbiosi dimostra una vitalità ed una resistenza superiore a quella delle due specie componenti isolate, ma non solo, il lichene possiede caratteri nuovi e costanti. Una data specie di fungo può entrare in simbiosi con diverse specie di alghe ed ogni associazione dà vita ad una diversa specie di lichene; allo stesso modo una data specie di alga può entrare in simbiosi con date specie di funghi creando diverse specie di licheni.
Un organismo lichenico possiede le caratteristiche della “specie biologica” è quindi una unità sistematica che può essere classificata.
Il fungo e l’alga se isolati dal lichene possono vivere separati solo su opportuni terreni di coltura, ma anche liberi in natura, esprimendo, il fungo, maggiori difficoltà nella sopravvivenza rispetto l’alga, ciò dimostra che il fungo ha subito maggiori processi di adattamento alla simbiosi.
La morfologia esterna dei licheni è molto varia, ma presto distinguibile da quella di altri esseri viventi, il corpo vegetativo è detto tallo ed è piuttosto omogeneo.
Rispetto la struttura interna è possibile distinguere talli omomeri e talli eteromeri. Nei primi le alghe sono distribuite uniformemente nella massa delle ife, nei secondi possono essere distinti degli strati: cortex superiore costituita da ife fortemente intrecciate, strato gonidiale che è frammisto ad ife lasse, medulla ad ife molto lasse, infine può essere presente una cortex inferiore provvista o meno di strutture di ancoraggio al substrato, le rizine.
ciglia, peli, pseudocifelle (con funzione di accrescere la superfici assorbente e di facilitare gli scambi gassosi alla superficie).
La riproduzione nei licheni può avvenire per via vegetativa e per via sessuale. La prima è affidata agli isidi ed ai soredi, estroflessioni superficiali del tallo contenenti ife e gonidi, che staccandosi dal tallo generante ricostituiscono un nuovo lichene clone. La seconda è affidata agli apoteci, forma molto comune di struttura riproduttiva, all’interno dei quali gli ascogoni producono gli aschi che generano le spore del fungo.
Gli apoteci hanno forma discoidale, la porzione superiore piatta e fertile è detta imenio; di forma globulosa sono i periteci che la cui porzione fertile comunica con l’esterno solo attraverso un poro detto ostiolo; di forma allungata o stellata è la lirella.
6.1 Storia dell’uso dei licheni.
Relativamente all’uso dei licheni (Richardson, 1974) se ne trova traccia già ai tempi dell’antico Egitto, dove Pseudoevernia furfuracea era una delle componenti usate per mummificare le salme; nell’antica Grecia ne erano conosciute le qualit{ coloranti, difatti prima dell’invenzione dei coloranti sintetici i licheni ebbero una grande importanza economica nella colorazione della lana; in Scandinavia Cetraria islandica veniva utilizzata nelle zuppe e nei biscotti; nella cosmesi è conosciuta l’essenza “muschio di quercia” che deriva da Evernia prunastri; nella farmacopea del nord Europa, tra il ‘500 e il ‘600, i licheni vengono apprezzati per le proprietà tonificanti e stimolanti; oggi ne è riconosciuto l’uso come antibiotici (es.:farmaci come Evosin e Usniplant sono ricavati da Parmelie e Usnee).
Recentemente è stato sintetizzato da talli di Umbelicariacee un carboidrato che ha mostrato una notevole attivit{ inibitrice nei confronti del virus dell’HIV.
Attualmente l’uso più interessante dei licheni riguarda la bioindicazione. Gi{ nel 1866 Nylander, in occasione di uno studio dei giardini del Lussemburgo, notò che i licheni presenti mostravano delle alterazioni morfologiche che egli collegò alle sostanze inquinanti presenti in atmosfera. Trent’anni dopo l’abate Huè verificò che l’intuizione di Nylander era corretta, i licheni di cui questi aveva parlato non esistevano più. Nel 1959 Grindon nel Lancashire meridionale osservò la sensibilit{ dei licheni all’inquinamento atmosferico. Nel 1969 Barkman propose per primo l’idea di utilizzare i licheni corticicoli per la valutazione della qualit{ dell’aria nei centri urbani e così via fino al 1974, anno in cui Ellenberg impostò in maniera rigorosa lo studio degli indicatori estendendolo dai vegetali superiori ai muschi e ai licheni. Ellenberg propose il concetto di bioindicazione come un insieme di valori assegnati a ciascuna specie vegetale, i quali ne quantificano il carattere di indicatore ambientale. In Italia dagli anni ’80 in poi grazie agli studi di Nimis e altri, inizia a diffondersi questa metodica che vede i licheni sempre più protagonisti nelle indagini di biomonitoraggio.
La grande sensibilit{ dei licheni all’inquinamento dell’aria è legata alla loro biologia; molte specie vivono per decine o centinaia di anni, questo li rende, come accade per le perennanti, soggetti all’effetto cumulativo degli inquinanti.
- la mancanza di meccanismi di escrezione attiva, quindi non possono “liberarsi” delle parti vecchie o danneggiate;
- la longevit{ e lentezza di accrescimento che permette di “leggere” sui talli lichenici gli effetti di prolungati periodi di esposizione a sostanze inquinanti.
Per determinate sostanze, come i metalli pesanti, gli idrocarburi clorurati, i radionuclidi, i floruri, i talli sono molto tolleranti, possono quindi assorbirne grandi quantità senza essere danneggiati, in questi casi sono ottimi bioaccumulatori2.
Sono invece molto sensibili alla presenza di altre sostanze come l’anidride solforosa, gli idrocarburi, l’ozono, gli ossidi di azoto, il nitrato di paracetile, il piombo, lo zinco, il cadmio, sensibilità che è stata accertata in esperimenti a fumigazione ed esposizione controllata; in ultimo ma non meno importante specie diverse hanno sensibilità diverse alle sostanze contaminanti.
La risposta fisiologica agli effetti dell’inquinamento atmosferico si può riassumere nelle seguenti reazioni:
- riduzione della fotosintesi per danneggiamento della clorofilla; (il principale inquinante responsabile è l’anidride solforosa);
- alterazione della forma e del colore del tallo;
- riduzione della fertilità, (rarefazione e riduzione delle dimensioni degli apoteci);
- alterazione delle comunità licheniche e diminuzione della copertura, (ad esempio è possibile osservare che nei centri cittadini le specie crostose prevalgono sulle fogliose fino alla totale scomparsa);
- riduzione del numero totale di specie nel tempo; - riduzione del numero totale di specie nello spazio.
2 Diploschistes muscorum può tollerare concentrazioni di metalli pesanti fino a 10 volti superiori a quelle riscontrate in natura (Sarret et al., 1998).
6.2 Il declino delle comunità licheniche.
E’ evidente che la distruzione degli habitat e degli ecosistemi su scala globale ha portato ad un deterioramento della flora lichenica, alla perdita di taxa e più in generale di biodiversità.
Deforestazione, pratiche agricole e inquinamento atmosferico hanno contribuito pesantemente al declino della flora lichenica (Coppins et. al., 2001; Nimis et al., 2002). E’ quindi urgente promuovere delle azioni di conservazione per il mantenimento della biodiversità e per fare questo è necessario conoscere la distribuzione e la presenza delle specie nei diversi territori (Sérusiaux, 1989). Come già accennato, le comunità licheniche sono oggi in regressione, la causa principale è certamente l’inquinamento atmosferico, ma recenti studi hanno messo in evidenza come le pratiche forestali giochino un ruolo importante sia nel mantenimento che nella perdita delle comunità. In particolare esse danneggiano le comunità a cianolicheni, che è risaputo sono indicatori di continuità forestale (Pykälä, 2006; Friedel et al., 2006; Nascimbene et al., 2007).
A sostegno di quanto detto è stato osservato che nei boschi cedui le comunità licheniche sono povere, cosa dovuta al fatto che con la ceduazione vengono materialmente rimossi insieme ai tronchi e ai rami, i licheni epifiti. All’azione di rimozione va aggiunto il drastico cambiamento relativo a luce ed umidità che si verifica nelle adiacenze degli alberi rimanenti che altera le condizioni ecologiche precedenti.
L’effetto finale è la distruzione della continuit{ ecologica anche nei siti che sono stati boscati per molto tempo (Rose & Coppins, 2002).
Ad esempio se consideriamo specie come Lobaria pulmonaria e Fuscopannaria
mediterranea che si riproducono per propaguli vegetativi con un efficacia di dispersione
piuttosto limitata, si comprende come interventi di taglio o di rimozione non selezionati ne danneggino in modo sensibile la presenza.
- I licheni sono importanti componenti della biosfera.
In casi particolari come nella tundra, alle alte latitudini, nei sistemi dunali, nei deserti come anche nelle aree urbane essi sono importanti nella catena alimentare; inoltre le comunità che si sviluppano e si succedono sulle rocce contribuiscono alla formazione del suolo (biodeteriogeni).
Conoscere, mantenere e promuovere attività di ricerca che possano implementare le azioni di conservazionismo nei confronti dei licheni significa proteggere anche il sistema che si evolve insieme agli stessi, sia che essi offrano rifugio e riparo alla fauna inferiore, sia che costituiscano la materia prima nelle complesse costruzioni dei nidi dei volatili come nelle tane di alcune specie di Aracnidi, sia che siano semplicemente nutrimento.
7 Materiali e metodi.
La ricerca è iniziata nella primavera del 2008. Sono stati scelti 18 plot in totale, all’interno dei quali sono stati raccolti i licheni e sono stati effettuati i rilievi per la biodiversità lichenica.
La scelta dei plot è avvenuta procedendo dalla cartografia cartacea alle immagini digitali e successivamente effettuando diversi sopralluoghi al fine di verificare le scelte attuate.
Per ogni area di saggio di dimensioni di 30mx30m sono state annotate le coordinate GPS in accordo col sistema italiano metodo Gauss-Boaga e l’altezza sul livello del mare.
7.1 Metodo di campionamento.
I licheni raccolti all’interno dei plot, su diversi tipi di substrato, sono stati identificati presso il laboratorio del DACPA usando uno steromicroscopio con ingrandimento fino a 50x ed un microscopio con ingrandimento fino a 65x, per poter riconoscere e misurare i caratteri utili all’identificazione. Molti dei talli raccolti sono stati esaminati dopo l’utilizzo di reattivi (K, Cl, KCl).
Sono state utilizzate per la determinazione dei campioni i testi seguenti: Clauzade & Roux (1985), Nimis (1987, 1991), Ravel (1997), Purvis et al., (1992), Bartholmess et
al., (2004), Sérusiaux et al. (2004), Wirth et al., (2004).
Per ogni plot, in base agli indici ecologici di Wirth, sono stati costruiti gli l’ecogrammi relativi agli indici di xerofitismo, igrofitismo e nitrofitismo; questo tipo di grafico permette in un colpo d’occhio di stimare le variabili ecologiche considerate.
Per il calcolo della biodiversit{ lichenica sono stati individuati all’interno dei plot, secondo la metodologia standard, tre forofiti rispondenti alle seguenti caratteristiche (Manuale ANPA, 2001): appartenere alla stessa specie arborea o per lo meno alla stessa classe (acidità/basicità della corteccia), avere una circonferenza minima di 60 cm, un’inclinazione minima del tronco di 10° e una bassa copertura muscinale, non essere alberi isolati o al margine dei boschi.
Sui forofiti prescelti è stato quindi apposto il reticolo3 composto da 4 subunità di
Ciascuna subunit{ è stata apposta sul tronco ai 4 punti cardinali ad un’altezza di 100 cm da terra (Fig. 17).
Sono state riempite le schede di rilevamento nelle quali vengono inseriti sia i dati relativi all’albero prescelto che tutte le specie presenti dentro le maglie del reticolo (Fig. 18). La frequenza dei licheni che può variare da 0 a 20, viene annotata per ogni maglia; la somma delle frequenze per ogni albero costituisce l’Indice di Biodiversit{ Lichenica (I.B.L.) della stazione (plot), (ANPA, 2001, 2003; Asta et al., 2002).
8 La flora lichenica dei SIC
8.1 Elenco floristico dei licheni epifiti del SIC “Bosco del Flascio”.
Anaptychia ciliaris (L.) Körb. Arthonia radiata (Pers.) Ach. Bacidia rosella (Pers.) De Not. Bacidia rubella (Hoffm.) A.Massal.
Caloplaca flavorubescens (Huds.) J.R.Laundon Caloplaca haematites (St.-Amans) Zwackh Caloplaca herbidella (Hue) H.Magn.
Caloplaca pyracea (Ach.) Th.Fr.
Candelariella xanthostigma (Ach.) Lettau Catillaria nigroclavata (Nyl.) Schuler Cladonia fimbriata (L.) Fr.
Cladonia pyxidata (L.) Hoffm. Collema flaccidum (Ach.) Ach.
Collema furfuraceum (Arnold) Du Rietz Collema nigrescens (Huds.) DC.
Degelia plumbea (Lightf.) M.Jørg. & P.James Diplotomma alboatrum (Hoffm.) Flot. Evernia prunastri (L.) Ach.
Fuscopannaria mediterranea (Tav.) M.Jørg.
Hyperphyscia adglutinata (Flörke) H.Mayrhofer & Poelt Hypogymnia physodes (L.) Nyl.
Hypogymnia tubulosa (Schaer.) Hav. Lecania fuscella (Schaer.) A.Massal.
Lecania naegelii (Hepp) Diederich & Van den Boom Lecanora argentata (Ach.) Malme
Lecanora carpinea (L.) Vain. Lecanora chlarotera Nyl. Lecanora hagenii (Ach.) Ach.
Lobaria pulmonaria (L.) Hoffm.
Melanelixia fuliginosa (Duby) O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. & Lumbsch Melanelixia glabra (Schaer.) O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. & Lumbsch Melanelixia subaurifera (Nyl.) O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. & Lumbsch Melanohalea elegantula (Zahlbr.) O.Blanco, A.Crespo, Divakar, Essl., D.Hawksw. &
Lumbsch
Melanohalea exasperata (De Not.) O.Blanco, A.Crespo, Divakar, Essl., D.Hawksw. &
Lumbsch
Nephroma bellum (Spreng.) Tuck. Ochrolechia balcanica Verseghy Parmelia saxatilis (L.) Ach. Parmelia sulcata Taylor
Parmelina tiliacea (Hoffm.) Hale Parmelina pastillifera (Harm.) Hale Parmelina quercina (Willd.) Hale Peltigera praetextata (Sommerf.) Zopf
Pertusaria albescens (Huds.) M.Choisy & Werner Pertusaria amara (Ach.) Nyl
Pertusaria coccodes (Ach.) Nyl. Pertusaria flavida (DC.) J.R.Laundon
Pertusaria hemisphaerica (Flörke) Erichsen Pertusaria hymenea (Ach.) Schaer.
Pertusaria leioplaca DC.
Pertusaria pertusa (Weigel) Tuck.
Phaeophyscia orbicularis (Neck.) Moberg Phlyctis argena (Spreng.) Flot.
Physcia adscendens (Fr.) H.Olivier Physcia aipolia (Humb.) Fürnrh. Physcia leptalea (Ach.) DC. Physcia tenella (Scop.) DC.
Physconia distorta (With.) J.R.Laundon Physconia perisidiosa (Erichsen) Moberg Physconia venusta (Ach.) Poelt
Pseudevernia furfuracea (L.) Zopf v. furfuracea Ramalina calicaris (L.) Fr.
Ramalina farinacea (L.) Ach. Ramalina fastigiata (Pers.) Ach. Ramalina fraxinea (L.) Ach. Ramalina roesleri (Schaer.) Hue Ramalina subgeniculata Nyl. Rinodina pyrina (Ach.) Arnold Xanthoria parietina (L.) Th.Fr.
8.2 Elenco floristico dei licheni epifiti del SIC “Rocche di Roccella Valdemone”.
Amandinea punctata (Hoffm.) Coppins & Scheid. Anaptychia ciliaris (L.) Körb.
Bacidia circumspecta (Vain.) Malme Bacidia rubella (Hoffm.) A.Massal. Caloplaca cerina (Hedw.) Th.Fr. v. cerina
Caloplaca cerinelloides (Erichsen) Poelt
Caloplaca ferruginea (Huds.) Th.Fr.
Caloplaca flavorubescens (Huds.) J.R.Laundon Caloplaca haematites (St.Amans) Zwackh Caloplaca herbidella (Hue) H.Magn. Caloplaca pyracea (Ach.) Th.Fr.
Candelariella xanthostigma (Ach.) Lettau Catillaria nigroclavata (Nyl.) Schuler Cladonia pyxidata (L.) Hoffm.
Collema nigrescens (Huds.) DC. Collema subflaccidum Degel. Collema subnigrescens Degel.
Diplotomma alboatrum (Hoffm.) Flot. Evernia prunastri (L.) Ach.
Hypogymnia tubulosa (Schaer.) Hav. Lecania cyrtella (Ach.) Th.Fr.
Lecania fuscella (Schaer.) A.Massal.
Lecania naegelii (Hepp) Diederich & Van den Boom Lecanora carpinea (L.) Vain.
Lecanora chlarotera Nyl. Lecanora hagenii (Ach.) Ach. Lecanora horiza (Ach.) Linds. Lecanora pulicaris (Pers.) Ach.
Lecidella elaeochroma (Ach.) M.Choisy Lepraria nivalis J.R. Laundon
Megaspora verrucosa v. mutabilis (Ach.) Nimis & Cl.Roux
Melanelixia fuliginosa (Duby) O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. & Lumbsch Melanelixia glabra (Schaer.) O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. & Lumbsch Melanelixia subaurifera (Nyl.) O. Blanco, A. Crespo, Divakar, Essl., D. Hawksw. & Lumbsch Melanohalea elegantula (Zahlbr.) O.Blanco, A.Crespo, Divakar, Essl., D.Hawksw. &
Lumbsch
Melanohalea exasperata (De Not.) O.Blanco, A.Crespo, Divakar, Essl., D.Hawksw. &
Lumbsch
Ochrolechia balcanica Verseghy Parmelia saxatilis (L.) Ach. Parmelia sulcata Taylor
Parmelina tiliacea (Hoffm.) Hale Parmelina quercina (Willd.) Hale Peltigera praetextata (Sommerf.) Zopf
Pertusaria albescens (Huds.) M.Choisy & Werner Pertusaria amara (Ach.) Nyl
Pertusaria flavida (DC.) J.R.Laundon
Pertusaria hemisphaerica (Flörke) Erichsen Pertusaria hymenea (Ach.) Schaer.
Pertusaria leioplaca DC.
Pertusaria pertusa (Weigel) Tuck. Phaeophyscia ciliata (Hoffm.) Moberg Phaeophyscia orbicularis (Neck.) Moberg Phlyctis argena (Spreng.) Flot.
Physcia adscendens (Fr.) H.Olivier Physcia aipolia (Humb.) Fürnrh.
Physcia biziana (A.Massal.) Zahlbr. v. biziana Physcia leptalea (Ach.) DC.
Physcia stellaris (L.) Nyl. Physcia tenella (Scop.) DC.
Pleurosticta acetabulum (Neck.) Elix & Lumbsch Pseudevernia furfuracea (L.) Zopf v. furfuracea Ramalina calicaris (L.) Fr.
Ramalina farinacea (L.) Ach. Ramalina fastigiata (Pers.) Ach. Ramalina fraxinea (L.) Ach. Ramalina subgeniculata Nyl. Rinodina colobina (Ach.) Th.Fr. Rinodina pyrina (Ach.) Arnold Rinodina sophodes (Ach.) A.Massal.
Tephromela atra v. torulosa (Flot.) Hafellner Xanthoria parietina (L.) Th.Fr.
8.3 Commento alla flora.
La ricerca ha portato al censimento di una flora alquanto ricca: 100 specie sono state ritrovate nel SIC Bosco del Flascio e 98 nel SIC Rocche di Roccella Valdemone. La nomenclatura è conforme alla check-list dei licheni d’Italia (Nimis & Martellos, 2008a). I licheni collezionati sono stati identificati dall’autore con l’aiuto del tutor e sono conservati in un erbario personale.
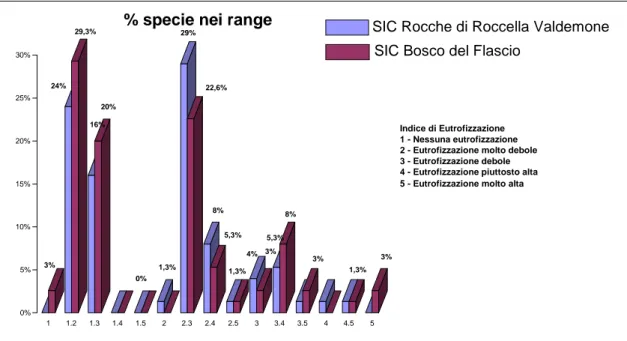
In particolare per l’analisi ecologica della flora (Fig. 19) sono stati considerati i licheni epifiti. Nel SIC Bosco del Flascio sono state ritrovate 72 entità epifite (71 specie, 1 varietà), di cui il 42% fogliose, il 14% fruticose, il 40% crostose, 1,30% squamolose, 1,30% leprose; nel SIC Rocche di Roccella Valdemone 75 (71 specie, 4 varietà) di cui il 38% fogliose, il 10% fruticose, il 48% crostose, 1,30% squamolose, 1,30% leprose.
Si sottolinea che nelle trattazioni seguenti si fa riferimento solo alla flora epifita. Le specie censite, seguendo http://www.biolib.cz/en/main/, appartengono a 16 famiglie e 38 generi, come di seguito riportato:
Arthoniaceae: Arthonia, Candelariaceae: Candelariella, Catillariaceae: Catillaria, Cladoniaceae: Cladonia,
Collemataceae: Collema, Leptogium, Lecanoraceae: Lecanora, Lecidella, Pannariaceae: Degelia, Fuscopannaria,
Parmeliaceae: Evernia, Hypogmnia, Melanelixia, Melanohalea, Parmelia, Parmelina,
Pleurosticta, Pseudoevernia,
Ramalinaceae: Bacidia, Ramalina, Tephromela, Lobariaceae: Lobaria,
Nephromataceae: Nephroma, Peltigeraceae: Peltigera,
Pertusariaceae: Ochrolechia, Pertusaria,
Physciaceae: Amandinea, Anaptychia, Diplotomma, Hyperphyscia, Phaeophyscia,
Physcia,Physconia, Rinodina.
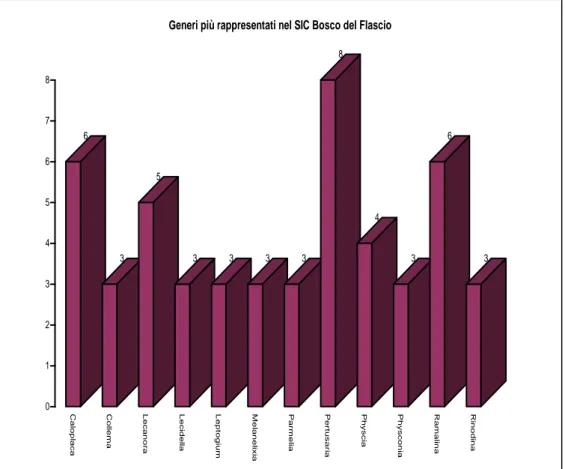
Si è misurata l’incidenza percentuale dei generi della flora epifita e sono stati messi in evidenza i generi maggiormente rappresentati (Figg. 20-21): nel SIC Rocche di Roccella Valdemone i generi più ricchi di specie sono Caloplaca e Pertusaria con 8 specie, seguite da Lecanora e Physcia con 6; nel SIC Bosco del Flascio il genere Pertusaria è presente con 8 specie, seguito dai generi Caloplaca e Ramalina con 6 (Figg.22-23).
21% 26% 17% 16% 48% 40% 10% 14% 1,30% 1,30% 0% 5% 10% 15% 20% 25% 30% 35% 40% 45% 50%
l.l. l.s cros frut squam lepr.
Confronto % forme di crescita
SIC Rocche di Roccella Valdemone
SIC Bosco del Flascio
Fig. 21
Incidenza % dei generi nella flora epifita del SIC Rocche di Rocche di Roccella Valdemone
0% 2% 4% 6% 8% 10% 12% 14% 16% 18% 20% Amandinea Anaptychia Bacidia Caloplaca Candelariella Catillaria Cladonia Collema Diplotomma Evernia Hyperphyscia Hypogymnia Lecania Lecanora Lecidella Lepraria Leptogium Megaspora Melanelixia Melanohaela Ochrolechia Parmelia Pamelina Peltigera Pertusaria Phaeophyscia Phlyctis Physcia Physconia Pleurosticta Pseudoevernia Ramalina Rinodina Xanthoria
Incidenza % dei generi nel SIC Rocche di Roccella Valdemone
Fig.20
Incidenza % dei generi nella flora epifita del SIC Bsoco del Flascio
0% 5% 10% 15% 20% 25% Anaptychia Arthonia Bacidia Caloplaca Candelariella Catillaria Cladonia Collema Degelia Diplotomma Evernia Fuscopannari Hyperphyscia Hypogymnia Lecania Lecanora Lecidella Lepraria Lobaria Melanelixia Melanohaela Nephroma Ochrolechia Parmelia Pamelina Peltigera Pertusaria Phaeophyscia Phlyctis Physcia Physconia Pleurosticta Pseudoevernia Ramalina Rinodina Xanthoria
8 4 4 6 3 3 8 6 4 5 3 0 1 2 3 4 5 6 7 8 Caloplac a C o lle ma L e ca n ia L e ca n o ra Me la n e lixi a Pe lt ig e ra Pe rt u sa ri a Ph ysci a Ph ysco n ia R a ma lin a Rinodina
Generi più rappresentati nel SIC Rocche di Roccella Valdemone
Fig. 23 6 3 5 3 3 3 3 8 4 3 6 3 0 1 2 3 4 5 6 7 8 C a lo p la ca C o lle m a L e ca n o r a L e ci d e lla L e p to g iu m M e la n e lixi a P a r m e lia P e r tu sa r ia P h ysci a P h ysco n ia R a m a lin a R in o d in a
Generi più rappresentati nel SIC Bosco del Flascio
8.4 Distribuzione delle specie in base ai piani fitoclimatici.
Si è ritenuto interessante confrontare la percentuale di specie reperite relativamente all’appartenenza al piano fitoclimatico (Fig. 24).
Nei grafici di Fig. 25 e 26 sono riportate le percentuali relative, le quali indicano una buona relazione tra piani fitoclimatici e bioclima: la distribuzione delle specie rispecchia il bioclima.
L’andamento delle piogge che per il SIC Bosco del Flascio, si assesta tra 800 e 1000 mm annui, ed è conforme col dato relativo alla presenza di specie proprie del piano Mediterraneo umido (67%); per il SIC Rocche di Roccella Valdemone il cui regime di piovosità va dai 1000 ai 1.300 mm annui le specie del piano Mediterraneo umido rappresentano l’85% coerentemente quindi al dato di piovosit{.
Fig.24: Il Paese suddiviso nei nove piani fitoclimatici (da Italic)
Fig.: 25 e 26 Ripartizione percentuale dei taxa
SIC Bosco del Flascio
1% 8% 67% 24% Montano Mediterraneo secco Mediterraneo umido SubMediterraneo umido
SIC Rocche di Roccella Valdemone 1% 3% 85% 11% Montano Mediterraneo secco Mediterraneo umido SubMediterraneo umido
9 Il concetto di autoecologia.
L’autoecologia è un indirizzo dell’ecologia che si occupa di studiare le relazioni tra gli organismi biologici e gli elementi non biologici in un determinato ambiente, essa analizza i rapporti con i vari fattori ecologici utilizzando metodi e tecniche delle scienze sperimentali. L’autoecologia studia la distribuzione delle singole specie cercando di isolare tutti i fattori ecologici che intervengono sulla loro stessa distribuzione.
Considerando gli organismi vegetali, essi ad esempio, durante il corso dell’evoluzione hanno dovuto sviluppare particolari adattamenti agli ambienti terrestri, dai temperati ai desertici sino a quelli salmastri, il che ha comportato il manifestarsi di cambiamenti che hanno coinvolto e coinvolgono sia i processi biologici interni che la morfologia esterna. Questi organismi hanno quindi necessità di bilanciare le proprie esigenze ecologiche in relazione alle variazioni dei fattori ambientali. Fondamentalmente si parla di tre tipi di bilancio:
- termico: modificazioni ed effetti sugli organismi vegetali in relazione alla temperatura e alla radiazione solare.
- idrico: strategie per mantenere nei tessuti e negli apparati vegetali un alto quantitativo di acqua.
- dei nutrienti: strategie per mantenere alto l’apporto dei nutrienti per la crescita degli organi vegetali.
Ogni volta che una pianta, per cause naturali, si trova a vegetare in un determinato sito, questo è una prova che il sito è compatibile con le sue esigenze ecologiche: dalla sua presenza si possono dunque ricavare informazioni sulle caratteristiche ecologiche del sito stesso.
Nel 1974 Ellenberg propose di utilizzare come metodo di bioindicazione l’enorme evidenza di dati raccolti sulla distribuzione geografica e topografica delle piante. La vegetazione può quindi essere interpretata come un segnale, che fornisce informazioni sulle condizioni ecologiche dell’ambiente (Pignatti, 1980). I valori di bioindicazione costituiscono la valutazione numerica del segnale che ciascuna specie fornisce, sull’incidenza dei principali fattori ecologici nel determinare le caratteristiche del sito: si tratta di una valutazione soggettiva, ma che tiene conto di una grande quantità di fatti obiettivi: distribuzione geografica e topografica della specie, misure sperimentali in campo, confronti con altre specie.
L’ambiente interviene sul vegetale “imponendo” le sue caratteristiche edifico-climatiche alle quali gli organismi rispondono principalmente in tre modi:
- allontanamento (la specie non si adatta alla stazione);
- tolleranza (la specie non scompare dalla stazione, ma presenta anche temporaneamente delle perfomance che le permettono di persistere anche se i fattori non gli sono idonei);
- adattamento (la specie presenta adattamenti al fattore sfavorevole fino ad arrivare al premio evolutivo).
9.1 Autoecologia dei licheni.
I licheni sono organismi ubiquisti pur essendo molto sensibili ai cambiamenti dell’ecosistema; questo loro successo è dovuto in gran parte alla grande capacit{ di adattamento alle variazioni di secchezza e umidit{: quando l’acqua viene a mancare essi smettono di crescere ed entrano in dormienza (Aubert et al., 2007) anche per lunghi periodi, alla presenza anche minima di acqua, grazie al fenomeno della reviviscenza, la assorbono fino ad imbibirsene 30 volte il loro peso riprendendo le funzioni vitali.
Essi colonizzano il legno e la scorza degli alberi, le rocce, il suolo, i muri, e i più diversi manufatti, ma non tutte le specie crescono sugli stessi substrati: ad esempio alcuni vivono sulle scorze ma non degli stessi alberi, poichè scorze di specie di alberi differenti presentano proprietà chimiche differenti, prima su tutte il pH, che creano condizioni ottimali solo per date specie.
Alcuni licheni crescono su scorze lisce e scompaiono quando con l’et{ la scorza diventa più rugosa; ancora alcune specie sono eliofile, altre sciafile, altre preferiscono i luoghi freschi e umidi, altre sono limitate agli habitat secchi e soleggiati. I fattori ambientali “distribuiscono” le comunit{ licheniche secondo gradienti che possono essere ben definiti. In tale senso è possibile distinguere macrotipi funzionali che danno informazioni relativamente al tipo di disturbo: i licheni a lobi stretti, ad alghe verdi e soredi (Physcia, Physconia, Phaeophyscia…), sono entità resistenti, si ritrovano infatti nei pascoli,negli arbusteti, nei seminativi, tutte aree queste dove possono verificarsi fenomeni di inaridimento con conseguente formazioni di polveri e d eutrofizzazione; licheni a tallo leproso, fruticoso o foglioso a cianobatteri, al contrario sonpo particolarmente sensibili ai disturbi ambientali e pertanto hanno il loro optimum negli habitat preservati.
Le comunità licheniche epifite in particolare, si distribuiscono principalmente secondo alcuni fattori quali la texture della corteccia, il pH, la capacità di ritenzione idrica, l’irradiazione solare, l’umidit{ e il grado di eutrofizzazione (nitrofitismo), colonizzando dai boschi naturali e seminaturali ricchi di specie licheniche diverse al deserto lichenico delle nostre città.
9.2 I fotobionti, eccetto i cianobatteri.
La maggior parte delle specie licheniche contiene come fotobionte alghe verdi (85%). Esse sono afferenti principalmente al genere
Trebouxia (Fig.29); poche contengono alghe
del genere Trenthepohlia (Fig.30), che sono molto comuni nei climi caldo umidi tropicali (Frield & Budel, 1996) e che colonizzano alle nostre latitudini siti particolari, come fondo cava e boschi umidi e ombreggiati. Fanno parte della flora italiana i licheni, a
Trenthepohlia, dei generi Arthonia,
Schimmatomma, Graphis, Ophegrapha. I
licheni a Trebouxia decisamente i più comuni, dominano dove le condizioni ambientali sono più aride e disturbate. La flora lichenica epifita censita nel SIC Bosco del Flascio, consta di 72 entità di cui 65 hanno come partner foto sintetico alghe verdi.
Una sola specie, Arthonia radiata ha come partner fotosintetico alghe del genere
Trentepohlia.
Per quel che riguarda la flora epifita censita nel SIC Rocche di Roccella Valdemone, essa consta di 75 entit{ di cui 71 sono licheni il cui fotobionte è un’alga verde.
Fig. 29: Alghe del genere Trebouxia
Fig.31: Alga del tipo Nostoc 9.3 I cianobatteri.
Quando il partner fotosintetico è un cianobatterio (Nostoc, Fig: 31 o altro genere) i licheni vengono chiamati cianolicheni (es.
Collema, Leptogium). In alcuni generi a Trebouxia sono presenti delle strutture nel
tallo dette cefalodi, in cui sono alloggiati i cianobatteri che costituiscono il partner fotosintetico secondario (Lobaria, Peltigera). In entrambe i casi i cianobatteri fissano azoto atmosferico, peculiare il fatto che siano rivestiti di mucillaggine la quale gioca un ruolo importante nel bilancio idrico del lichene.
Dei licheni conosciuti solo il 10% sono cianolicheni; ma localmente dove le condizioni ecologiche sono favorevoli, (es. habitat maturi, vecchie foreste o foreste oceaniche), essi possono essere molto abbondanti.4 In accordo con Rose (1988) è
possibile affermare che i generi che comprendono cianobatteri come fotobionte, tipo
Collema, Leptogium, Nephroma, Pannaria, Peltigera, appartengono alle comunità del Lobarion pulmonariae, Ochsner 1928, le quali erano molto diffuse in tutta Europa fin
nelle pianure ed il cui declino è da attribuirsi all’’inquinamento atmosferico da acidificanti (Gries, 1996; Wirth, 1988) ed anche a determinate pratiche forestali (Richardson & Cameron 2004) che ne limitano la dispersione (Pykälä, 2004).
I cianolicheni possono essere in genere considerati non comuni o rari (Rabinowitz, 1981), pertanto all’interno di regioni geografiche può verificarsi una delle tre condizioni:
- elevata abbondanza in poche località;
- bassa abbondanza in un largo numero di località; - bassa abbondanza in poche località.
E’ possibile supporre che soprattutto nel Bosco del Flascio il numero di cianolicheni sia più elevato trattandosi di un habitat ben conservato e con un microclima più umido dovuto alla presenza del fiume.
4 In alcuni casi possono anche eccedere i licheni ad alghe verdi, come è il caso delle foreste di cedro e abete canadese della Columbia Britannica (Cameron et al, 2006).
Relativamente alla scala di cui sopra, si può dire che il SIC Bosco del Flascio si trovi nella II condizione; nella III il SIC Rocche di Roccella Valdemone.
Nelle pagine seguenti vengono considerati solo i licheni epifiti poiché più sensibili all’inquinamento dell’aria, rispetto a quelli epilitici e terricoli (Richardson & Cameron l.c.).
Nel SIC Bosco del Flascio si evidenziano 8 specie; 5 specie nel SIC Rocche di Roccella Valdemone.
9.4 I cianolicheni del SIC Bosco del Flascio.
Collema Wigg.
Collema flaccidum (Ach) Ach.: è un lichene molto diffuso dalla regione artico
boreale fino alla regione mediterranea, molto frequente nelle aree a clima oceanico. E’ più facile rinvenirlo su rocce silicee umide e poste in ombra, ma è possibile ritrovarlo anche sulle scorze.
E’ stato ritrovato in tutta Italia dove è piuttosto comune nelle aree umide, ma nelle zone in cui l’impatto antropico è veramente pesante è del tutto scomparso -ad esempio nella pianura padana- (Nimis & Tretiach, 1999).
Collema furfuraceum (Arnold) Du Rietz: molto diffuso in Europa dalla regione
boreale a quella mediterranea.
Cresce di solito su scorza di alberi vecchi in boschi aperti o su alberi isolati spesso associato a Nephroma, Leptogium e Lobaria; al sud è più facile ritrovarlo nei territori montani (Nimis, 1993a; Nimis & Tretiach, 1999).
Spesso confuso con Collema nigrescens in Italia è stato raramente campionato. Si rinviene anche all’interno di comunità licheniche nitrofiche e xerofitiche.
Collema nigrescens (Huds.) DC.: il suo areale di distribuzione coincide con quello
di C. furfuraceum.
Cresce su scorza acida o debolmente acida, su muschi epifitici e occasionalmente su roccia umida in aree dove le precipitazioni sono abbondanti.
Si ritrova sia nei boschi ben conservati che in zone piuttosto nitrofiche. In Italia è piuttosto comune nelle aree a clima caldo-umido (Nimis, 1993a).
Probabilmente la specie era molto più comune nel passato ma oggi è scomparsa nelle aree maggiormente disturbate (Nimis & Tretiach, 1999).
Degelia Arv. & D. J. Galloway
Degelia plumbea (Lightf.) M. JØrg. & P. James: in Europa è una specie
principalmente oceanica; nella regione Mediterranea si ritrova tra i 500 e i 900 m s.l.m.. Cresce su roccia, ma che non di rado colonizza la scorza degli alberi preferendo habitat umidi e ombreggiati.
In Italia è piuttosto rara, ma frequente nelle aree montane soprattutto a est della penisola (Nimis, 1993a).
Fuscopannaria P.M. JØrg.
Fuscopannaria mediterranea (Tav.) M. JØrg.: è questa una specie
Mediterraneo-Atlantica, molto frequente tra i 400 e i 900 m s.l.m. nella regione mediterranea.
Cresce principalmente nelle fessure della scorza, a volte tra i muschi in ambienti umidi, ma è stata ritrovata anche in condizioni di clima secco. In Italia è piuttosto rara, la si ritrova sopratutto lungo la costa Tirrenica (Nimis, 1993a).
Lobaria (Schreb.) Hoffm.
Lobaria pulmonaria (L.) Hoffm.: specie suboceanica, diffusa in Europa dalla
regione boreale a quella mediterranea (Purvis et al. 1992). Cresce in habitat con alta umidit{ atmosferica, su scorza acida di latifoglie; è molto sensibile all’inquinamento atmosferico e a pratiche di gestione forestale.
Essa è considerata da diversi autori (Rose, 1974, 1976, 1988; Gustafsson et al. 1992) un ottimo indicatore di continuità ecologica dei boschi; a causa del generale declino è stata inserita come specie vulnerabile nella Lista Rossa dei Licheni d’Italia (Nimis, 1992).
Nephroma Ach.
Nephroma bellum (Spreng.) Tuck.: questa specie è diffusa in Europa dalla regione
boreale a quella mediterranea (Wirth, 1995a).
In Italia è veramente rara e sensibile alla gestione forestale (Nimis, 1993a). La si trova su scorza o muschi epilitici del piano montano in zone molto piovose. In Italia è piuttosto rara.
Peltigera Willd.
Peltigera praetextata (Sommerf.) Zopf: è una specie comune nelle zone temperate
e sub-boreali d’Europa, principalmente nelle aree oceaniche (Vitikainen, 1994).
Nella regione mediterranea è la più comune Peltigera, la si ritrova su diversi substrati, su scorza, su ceppi, su roccia da basica ad acida, a volte anche al suolo (Martinez & Burgaz, 1997). In Italia è molto frequente e spesso confusa con Peltigera
9.5 I cianolicheni del SIC Rocche di Roccella Valdemone.
Collema Wigg.
Collema nigrescens (Huds.) DC. (cfr. sopra)
Collema subflaccidum Degel: è una specie ad ampia distribuzione in entrambi gli
emisferi (Nimis, 1993a). Cresce sulle scorze soprattutto di vecchi alberi, raramente la si ritrova su roccia (Purvis et al. 1992). In Italia è presente soprattutto al nord e lungo il Tirreno nelle zone a clima piuttosto umido (Nimis, 1993a). E’ scomparsa nei siti pesantemente disturbati (Nimis & Tretiach, 1999).
Collema subnigrescens Degel.: ha ecologia molto simile a quella di C. nigrescens,
preferisce però stazioni più igrofitiche e ombrofitiche. Cresce su scorze anche molto ricche in nutrienti e su roccia. In Italia può essere localmente frequente nelle aree a clima umido e in montagna; si è praticamente estinto dove c’è inquinamento atmosferico (Nimis & Tretiach, 1999).
Leptogium (Ach.) Gray
Leptogium teretiusculum (Wallr.) Arnold: è una specie propria di regioni
temperate.
Cresce specialmente sui muschi, su scorze rugose subneutre, di rado al suolo o su roccia (JØrgensen 1994, Wirth 1995a). In Italia sono state fatte poche segnalazioni, probabilmente per confusione altri Leptogium (Nimis, 1993a).
Peltigera Willd.
9.6 L’indice di Poleofobia (P).
Questo indice esprime la tendenza di una specie lichenica ad occupare aree interessate da differenti gradi di disturbo; esso è espresso da quattro livelli di cui il primo corrispondente a P=0 indica specie che crescono solo su vecchi alberi in boschi non disturbati. Questo livello è l’unico dei quattro che è riferito solo a specie epifite.
L’ultimo livello P=3 indica specie che possono vivere in ambienti disturbati comprese le città. I valori intermedi indicano situazioni intermedie.
P=1 specie che vivono in habitat naturali o seminaturali.
P=2 specie che vivono in aree moderatamente disturbate come aree agricole E’ da rilevare che tre specie del gruppo delle rare del SIC Bosco del Flascio hanno P=0: Bacidia rosella, Degelia plumbea e Nephroma bellum.
Per gli altri livelli di poleofobia (Fig.33) le specie sono così ripartite: P=1: 14 specie; P=1-2: 37 specie; P=1-3: 18specie.
Significativo il numero di specie con indice di poleofobia compresa tra P=1 e P=2, indicante che il 51% sul totale delle specie epifite predilige habitat seminaturali o moderatamente disturbati.
Anche per il SIC Rocche di Roccella Valdemone sono state evidenziati i livelli di poleofobia indicando che: P=0 nessuna specie; P=1: 12 specie, P=1-2: 44 specie; P=1-3: 19 specie.
È evidente la prevalenza di specie con indice di P=1-2, che rappresentano 58% del totale delle epifite (Fig. 32).
Questo indice congiunto all’indice di rarit{ e la presenza dei cianolicheni ci permette di asserire una interessante osservazione: oggettivamente ci troviamo di fronte due situazioni stazionali piuttosto simili ma che a questo punto dimostrano come le norme di salvaguardia e il miglior stato di conservazione del SIC Bosco del Flascio
0 4% 16% 19,5% 58% 51% 25% 22% 0% 10% 20% 30% 40% 50% 60% "Pl0" "Pl1" "Pl1,2" "Pl1,3"
Confronto % specie poleofobiche SIC Rocche di Roccella Valdemone SIC Bosco del Flascio
P=0 indica specie che crescono solo su vecchi alberi in boschi non disturbati. P=1 specie che vivono in habitat naturali o seminaturali
P=2 specie che vivono in aree moderatamente disturbate come aree agricole
P=3 indica specie che possono vivere in ambienti disturbati comprese le città.