Capitolo 1
Il senso del tatto
1.1 La pelle e la sua struttura:
La pelle (o cute) è il rivestimento esterno del corpo umano e rappresenta la prima interfaccia tra uomo ed ambiente che lo circonda.
Per le funzioni alle quali provvede e per la collocazione sul corpo, essa costituisce la prima e principale sorgente di ingresso sensoriale verso il sistema nervoso centrale e fornisce informazioni che riguardano sia l’ambiente esterno immediatamente prossimo, attraverso il contatto diretto con oggetti che lo compongono, sia sorgenti remote, attraverso la rilevazione della radiazione termica.
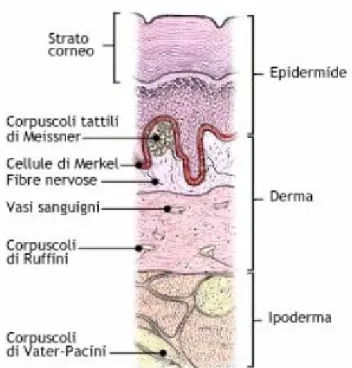
La cute è formata da tre strati principali: l’epidermide in superficie, il derma in posizione intermedia e l’ipoderma, lo strato più profondo che svolge funzioni di riserva energetica per l’organismo (Fig.1).
Fig. 1: Sezione trasversale della cute e collocazione delle cellule che la compongono.
L’epidermide è lo strato più esterno della cute, di spessore compreso tra 50 e 1,5 micron che dipende, per nutrizione e sensibilità, dal derma, in quanto ad essa non confluiscono né vasi sanguigni, né fibre nervose. Le cellule che la costituiscono sono morfologicamente differenti e formano cinque strati: lo strato basale, a contatto col derma da cui lo separa
una membrana, formato da cellule in grado di riprodursi, lo strato spinoso, immediatamente sopra, che svolge funzioni meccaniche, lo strato granuloso, che contribuisce a rendere l’epidermide impermeabile ed a conferirle le diverse caratteristiche protettive, lo strato lucido e lo strato corneo, a contatto con l’ambiente esterno, formato da cellule molto appiattite e sprovviste di nucleo.
Il derma è uno strato robusto ed elastico, di spessore medio compreso tra i 2 e 3 mm, in cui confluiscono numerosi vasi sanguigni e linfatici e una grande quantità di fibre nervose. Essa contiene i bulbi piliferi ed i recettori tattili.
La superficie totale cutanea nell’adulto, pur variando da individuo ad individuo in rapporto all’età, alla costituzione fisica ed al sesso è stimata intorno a 1,5 ; 1,8 mq, mentre il suo spessore medio, diverso nelle varie parti del corpo, è compreso tra 1,5 e 2 mm nel palmo delle mani e nella pianta del piede ed i 4 mm nel cuoio capelluto.
1.2 I recettori tattili:
La sensibilità tattile (o tatto-pressoria) risulta dall’integrazione di segnali provenienti da differenti tipi di recettori cutanei sensibili agli stimoli meccanici, ma aventi diverse caratteristiche e diversa ubicazione nello spessore della cute.
In base al tipo di sensazione percepita (meccanica, termica o di dolore), i recettori tattili vengono comunemente suddivisi in tre grandi classi: i meccanorecettori (o meccanocettori) che rispondono a sollecitazioni di tipo meccanico (pressioni, vibrazioni, cinestesia), i termorecettori (o termocettori) che rispondono a sollecitazioni di tipo termico e i nocicettori che rispondono a forti e potenzialmente nocive sollecitazioni ambientali sia meccaniche che termiche e sono i responsabili della percezione del dolore.
Per la realizzazione di dispositivi di ausilio per non vedenti, i canali percettivi che maggiormente interessano sono quelli stimolati da sollecitazioni di tipo meccanico la cui rivelazione compete ai meccanocettori e quelli stimolati da sollecitazioni di tipo termico che riguardano i termocettori. La percezione, infatti, di diverse sequenze codificate di simboli, lettere, numeri, note musicali attraverso specifiche sollecitazioni pressorie o vibratorie può permettere al non vedente di accedere alla scrittura, alla lettura, alla “visione tattile” di un’immagine. La percezione, invece, del flusso di energia termica risultante dai termocettori può essere utilizzata per aggiungere dettagli e caratteristiche qualitative all’immagine tattile, precedentemente rivelata nelle sue caratteristiche essenziali, grazie alla stimolazione dei meccanocettori.
1.2.1 Sensibilità meccanica: i meccanorecettori
I meccanocettori hanno un’alta sensitività alla penetrazione, alla pressione ed alla vibrazione sulla pelle o al movimento dei peli.
Questi recettori rispondono tutti alla deformazione meccanica ma mostrano un diverso grado di adattamento allo stimolo: quelli che si adattano più prontamente (recettori fasici) rispondono in gran parte a stimoli meccanici rapidamente variabili nel tempo (vibrazioni di frequenza elevata), mentre quelli con adattabilità minore rispondono invece anche a stimoli meccanici che variano lentamente (vibrazioni a bassa frequenza) o che siano continui (stimoli pressori) [1].
Sulla base delle differenti modalità di adattamento, i meccanocettori vengono suddivisi in due classi: recettori ad adattamento rapido (RA) e recettori ad adattamento lento (SA). I meccanocettori RA sono presenti sia nella pelle glabra che nella pelle con peli. Essi vengono fortemente sollecitati dal movimento della pelle e ne codificano la velocità nella frequenza di scarica. Se la pelle viene sollecitata con uno stimolo a rampa seguito da uno stimolo costante, essi rispondono esclusivamente quando esiste movimento (ovvero in corrispondenza del tratto variabile nel tempo dello stimolo) e non quando la pelle è mantenuta ferma nella sua nuova posizione (ovvero in corrispondenza del tratto costante dello stimolo).
Ad una velocità costante di penetrazione le unità afferenti RA scaricano un treno di impulsi a frequenza proporzionale alla velocità di spostamento.
I meccanocettori RA includono due sottoclassi di recettori RA: i recettori RA1 che rispondono più selettivamente a stimoli di velocità ed i recettori RA2 che reagiscono più selettivamente a stimoli di accelerazione.
Alla classe dei recettori RA2 appartengono i corpuscoli di Pacini, situati nel tessuto sottocutaneo e quindi nello strato più profondo della pelle; si tratta di recettori a prontissimo adattamento che rispondono selettivamente alla stimolazione vibratoria della pelle con frequenze anche molto elevate (oltre 50 e fino a 300 Hz).
La classe dei recettori RA1 comprende, invece, i seguenti corpuscoli sensitivi:
- I corpuscoli di Meissner che si trovano riuniti in gruppi di due o tre alla sommità delle papille del derma. Tali recettori posseggono un adattamento abbastanza rapido, per cui la loro scarica in risposta ad uno stimolo continuo, si esaurisce nel giro di qualche secondo dall’inizio della stimolazione, mentre il massimo della risposta si
ottiene applicando una stimolazione vibratoria che superi la frequenza di pochi cicli al secondo. Il range effettivo di frequenza di uno stimolo vibratorio percepibile da questo recettore è compreso tra i 10 e i 200 Hz. I corpuscoli di Meissner sono numerosi nella cute glabra, ma quasi assenti nella cute con peli, dove la loro funzione è praticamente assolta da terminazioni sensitive che prendono contatto con la radice dei peli e che sono note come recettori del follicolo pilifero;
- I corpuscoli di Krause che si credeva fossero recettori del freddo, ma che, invece, hanno tutte le caratteristiche di un recettore RA1: infatti risultano essere sensibili ad un range di frequenze superiore ai 10 – 100 Hz e non danno risposte ad una pressione mantenuta stabilmente sulla pelle.
I meccanocettori SA sono recettori di posizione che, come i rivelatori di velocità, rispondono in seguito ad un movimento della pelle, ma in aggiunta sono in grado di sostenere una scarica di impulsi anche quando la pelle è mantenuta ferma in una nuova posizione. A differenza dei recettori RA, i meccanocettori SA possono fornire informazioni circa i cambiamenti a lungo termine delle condizioni meccaniche della pelle.
Riconsiderando uno stimolo a rampa seguito da uno stimolo costante, si vede che sono presenti scariche neurali anche nella condizione di costanza della sollecitazione; durante il plateau della pressione la velocità di scarica diminuisce fino a raggiungere un valore caratteristico per ogni recettore SA proporzionale all’ammontare della pressione stessa. La frequenza della risposta dei recettori RA ad una deformazione mantenuta costante nel tempo può essere aumentata raffreddando la pelle. Tuttavia la risposta dinamica ha comunque durata di pochi secondi, a differenza di quanto avviene nei termocettori in cui la scarica permane per un tempo più lungo, dell’ordine del minuto.
Alla classe dei recettori SA appartengono i seguenti corpuscoli:
o Le cellule di Merkel, situate alla base dell’epidermide, che sono recettori tattili ad adattamento molto basso e quindi capaci di rivelare stimoli di lunga durata; o Le terminazioni di Ruffini, allocate negli strati profondi del derma, che sono
recettori tattili praticamente sprovvisti di adattamento e quindi in grado di rispondere a stimoli di lunghissima durata. Tali corpuscoli possono, dunque, fornire una continua indicazione dell’entità della pressione o della tensione permanente nella pelle, conferendole la sensibilità pressoria.
1.2.2 Sensibilità termica: i termocettori
I termocettori sono recettori tattili sensibili alle variazioni di temperatura (e insensibili a stimolazioni meccaniche) che generano una scarica continua di impulsi se la pelle è mantenuta a temperatura costante, mentre producono un incremento/decremento del numero di impulsi quando la temperatura varia [3].
Esistono due classi di termocettori: recettori del freddo con risposta massima nel range di temperature 25 – 30 °C e recettori del caldo sensibili fino a temperature massime dell’ordine dei 40 – 45 °C.
I recettori del caldo rispondono alla stimolazione termica con un aumento della frequenza di scarica all’aumentare della temperatura cutanea al di sopra dei 30 – 35 °C fino a circa 40 – 45 °C , limite oltre il quale la frequenza comincia a diminuire all’ulteriore aumento della temperatura. Viceversa i recettori del freddo aumentano la propria frequenza di scarica col diminuire della temperatura cutanea al di sotto di 30 – 35 °C fino a circa 20 – 25°C, limite sotto al quale la loro frequenza diminuisce all’ulteriore ridursi della temperatura. Questa alta sensitività dinamica permette ai termocettori di rispondere a piccole e lente variazioni (meno di 1°C in 30 s) nella temperatura delle pelle.
1.3 Processo di trasduzione e codificazione degli stimoli
nei recettori
Gli stimoli fisici che, attraverso la pelle, giungono ai recettori hanno diversa natura, tuttavia il sistema nervoso è in grado di fornire una sensazione corrispondente e specifica a tali stimoli. Il risultato dell’azione del sistema nervoso cutaneo è la percezione di sensazioni distinte quali pressione, vibrazione, caldo, freddo etc.
I recettori convertono tutte le informazioni contenute nei diversi tipi di stimoli in una scarica di potenziali d’azione inviati ai centri nervosi tramite le fibre nervose sensitive. Questa conversione avviene in due fasi successive [2]:
1. la trasduzione, che è la conversione dello stimolo in un segnale elettrico “analogico” in grado di attivare le terminazione nervose; essa avviene nella parte propriamente recettiva della struttura recettoriale. Il segnale elettrico che si rigenera con la trasduzione è detto potenziale del recettore. Esso è una variazione del potenziale di membrana che riproduce abbastanza fedelmente l’andamento
dello stimolo; di norma si tratta di una depolarizzazione della membrana recettiva della terminazione assonale o di quella apicale della cellula sensoriale. Il potenziale del recettore ha carattere locale (ossia è limitato all’area ove cade lo stimolo) ed è graduale e quindi ben diverso dal potenziale d’azione. La trasduzione è un processo attivo in cui si attua una notevolissima amplificazione del segnale, utilizzando energia di origine cellulare.
2. la codificazione, ossia la conversione del potenziale del recettore in una scarica di potenziali d’azione la cui frequenza dipende dall’intensità dello stimolo. Ciò avviene nella parte della struttura recettoriale che costituisce l’”encoder” del recettore.
I recenti studi sul meccanismo di trasduzione dell’informazione tattile sono focalizzati principalmente al modo in cui le informazioni provenienti dai recettori vengono integrate alla periferia ed alla maniera in cui le differenti informazioni vengono messe insieme per dar vita alla complessa percezione tattile.
1.4 Meccanismo di localizzazione dello stimolo tattile:
La sensazione finale, così come la percepiamo, va intesa come la risultante della stimolazione di gruppi di recettori tattili e non come effetto della sollecitazione del singolo recettore. Una stimolazione anche puntiforme coinvolge diverse unità sensoriali.
Si definisce campo recettivo di un meccanocettore la superficie, generalmente circolare o ellissoidale, all’interno della quale uno stimolo puntiforme di una specifica intensità può eccitare il recettore. Il campo recettivo del recettore varia in base alla posizione del corpo sul quale è ubicato il recettore stesso: le unità di stimolazione tattile delle dita di una mano hanno un campo recettivo di 1 – 2 mm . Johansson e Vallbo hanno stimato che la 2
stimolazione localizzata in 2 mm nel dito di una mano eccita circa 100 unità RA e 30 SA, mentre in 50 micron può eccitare 10 unità RA e nessuna SA.
A causa della diversa distribuzione nella pelle, la risoluzione spaziale, di circa 1 mm, che si riscontra nei polpastrelli della mano umana sembra coinvolgere soprattutto le unità SA e RA2 che sono quelle responsabili della rilevazione delle forme fini degli oggetti in contatto.
L’organizzazione dei recettori in unità sensoriali, cui corrispondono campi recettivi di una certa estensione, pone un limite alla precisione nel localizzare un particolare punto della pelle stimolato.
Sembra, pertanto, logico prevedere che due stimoli puntiformi che cadano nello stesso campo recettivo non possano essere discriminati spazialmente, perché la scarica recettoriale che essi producono viene convogliata da una stessa fibra nervosa afferente; in effetti il potere discriminativo spaziale, definito come la capacità di percepire come distinti due stimoli prodotti da due punte separate, della sensibilità tattile di una regione è tanto maggiore quanto più è elevata la densità dei rispettivi recettori e quanto più ristretti sono i loro campi recettivi. Al crescere numerico dei campi recettivi corrisponde, ovviamente, un proporzionale aumento numerico delle fibre nervose afferenti, cioè una maggior “ricchezza” dell’innervazione sensitiva di una determinata regione del corpo; d’altro canto, è regola generale per tutte le sensibilità che i campi recettivi, anziché affiancati uno all’altro, siano più o meno sovrapposti tra di loro con la loro parte periferica: una organizzazione che sembrerebbe diminuire la capacità di localizzazione sensoria, ma che in realtà permette di accrescere la precisione nell’individuazione del punto stimolato.
L’informazione, infatti, su ciascuno dei caratteri degli stimoli non viene trasmessa in modo separato su una propria fibra afferente, bensì utilizzando un gruppo di fibre nelle quali la frequenza della scarica afferente è distribuita secondo un “quadro” (“pattern code”) che differisce per ogni carattere dello stimolo [1].
E’ evidente che le informazioni circa l’intensità e la localizzazione dello stimolo potrebbero essere trasmesse senza problemi se ogni minima area della superficie cutanea spazialmente discriminabile col tatto fosse il campo recettivo di una singola unità sensoriale; in questo modo, dunque, la localizzazione dello stimolo verrebbe definita dall’unità sensoriale che viene attivata e l’intensità dello stimolo dalla frequenza della scarica che in essa origina.
Un’organizzazione di questo tipo, per raggiungere l’altissima capacità discriminativa tattile che possiedono molte regioni della superficie cutanea, richiederebbe un numero di campi recettivi e di fibre nervose afferenti altrettanto elevato per unità di superficie cutanea; è noto, invece, che i campi recettivi tattili sono molto più estesi della distanza minima discriminabile tattilmente.
C1 C2
Fig.2: Sovrapposizione tra le periferie recettive di 2 unità sensoriali.
Facendo riferimento alla Fig.2, si considera la situazione, vicina alla realtà, in cui due unità sensoriali (U1 e U2) si sovrappongono con la parte periferica dei loro campi recettivi (C1 e C2) e sono tributarie delle due fibre afferenti (f1 e f2).
Uno stimolo (S) di una certa intensità, applicato al centro di C1, darà una risposta di intensità pari ad S nella fibra f1 e una risposta nulla nella fibra f2; viceversa, lo stesso stimolo applicato al centro di C2 darà una risposta di intensità pari ad S nella fibra f2 e una risposta nulla nella fibra f1.
Intensità e localizzazione dello stimolo sono quindi ben definite e distinte: l’intensità dalla frequenza della scarica e la localizzazione dalla fibra che la convoglia.
La discriminazione spaziale del sistema delle due unità recettive è però ben maggiore della distanza tra i centri dei rispettivi campi recettivi; infatti, immaginando di spostare lo stimolo di intensità S gradualmente dal centro di C1 al centro di C2, la frequenza della scarica, in modo proporzionale allo spostamento, in f1 diminuirà da S a 0 ed in f2 aumenterà da 0 a S.
La localizzazione dello stimolo potrà allora essere definita “in continuo” per tutti i punti del percorso tra i centri di C1 e di C2: è immediato calcolare che il rapporto tra la frequenza delle due scariche in f1 e f2 sarà uguale al rapporto tra le distanze del punto ove cade lo stimolo rispettivamente dal centro di C1 ed il centro di C2.
Le due complete informazioni su intensità e localizzazione dello stimolo possono essere facilmente “estratte” dalle scariche delle due fibre dal semplice confronto delle rispettive frequenze.
informazioni in un ristretto numero di “canali” di trasmissione senza che ne vada perduta alcuna parte del contenuto [2].
La soglia di discriminazione spaziale, definita come la distanza tra due sollecitazioni puntiformi a partire dalla quale il soggetto non avverte più sensazioni distinte, viene determinata stimolando due punti della cute simultaneamente con un apposito compasso. Tale soglia risulta essere massima nella punta delle dita (2,3 mm) e minima nella coscia 67 mm).
Si osserva, inoltre, che se la stimolazione è accompagnata da un riscaldamento del punto di applicazione, la capacità di discriminazione aumenta.
1.5 Trasporto dell’informazione tattile:
I segnali inerenti alle diverse modalità sensoriali vengono portati dai recettori ai centri nervosi spinali ed encefalici da classi di fibre nervose afferenti che differiscono per diametro e per velocità di conduzione [1].
Nel risalire dal midollo spinale ai centri encefalici, le fibre sensitive che portano i segnali sensitivi al talamo ed alla corteccia celebrale, costituiscono due principali vie: quella dei cordoni dorsali (via lemniscale) e quella dei cordoni ventro-laterali (via spino-talamica). Le fibre della via lemniscale, per la maggior parte mieliniche di grande o medio diametro e con velocità di conduzione più elevata, convogliano la sensibilità tattile fine, vibratoria e quella cinestetica; si tratta, quindi, di sensibilità fini e discriminative.
Le fibre della via spino-talamica, di diametro minore con velocità di conduzione relativamente bassa, convogliano un ampio spettro di sensibilità: quella termica (caldo e freddo), quella tatto-pressoria grossolana, quella dolorifica, nonché alcune sensibilità particolari come il prurito; si tratta, quindi, di sensibilità più grossolane e meno discriminative di quelle trasmesse tramite la via lemniscale.
1.6 Aree sensitive corticali interessate:
Le vie sensitive che, decorrendo nei cordoni dorsali e ventro-laterali del midollo spinale (lemniscale e spino-talamica) raggiungono il nucleo del talamo, proiettano poi in gran parte, tramite neuroni talamo-corticali, all’area sensitiva primaria (SI) della corteccia celebrale; essa è situata posteriormente alla scissura centrale di Rolando.
Sollecitando una regione cutanea con uno stimolo tattile, si osserva una variazione di potenziale localizzata nella corrispondente regione dell’area SI; questo potenziale evocato si può derivare con un elettrodo posto sulla corteccia stessa o, più facilmente, sul cuoio
capelluto. Col metodo della derivazione dei potenziali evocati è possibile ottenere per la sensibilità tattile una mappa molto precisa delle localizzazioni corticali sensitive.
Esperimenti di derivazione dei potenziali corticali evocati da stimoli tattili e propriocettivi hanno dimostrato che i segnali tattili giungono prevalentemente alla parte anteriore dell’area SI, mentre quelli propriocettivi giungono alla parte posteriore; i due tipi di segnali interessano, perciò, almeno al loro arrivo, aggregati distinti di neuroni corticali.
Le proiezioni sensitive sull’area SI sono evidenti soprattutto per la sensibilità tattile fine, caratterizzata da un elevato potere discriminativo spaziale.
La corteccia della SI non è, comunque, un semplice luogo di proiezione, topograficamente ordinata, dei segnali sensitivi delle diverse parti del corpo. E’ noto, ad esempio, che le vie sensitive presentano, nel loro decorso dai recettori fino alla corteccia, una rilevante divergenza; in ciascuna delle stazioni disposte in catena nelle vie sensitive, ogni neurone che procede è connesso, tramite collaterali, con un numero crescente di neuroni che seguono. Ne deriva che una stimolazione cutanea, anche puntiforme, determina attivazione neuronica in un’area corticale molto estesa, nella quale i neuroni centrali sono più attivati di quelli periferici; inoltre la stimolazione di due punti vicini della superficie cutanea implica l’attività di due aree corticali ampiamente sovrapposte.
Una maggior estensione delle singole aree recettive corticali male si accorda con l’elevata precisione con cui le diverse parti del corpo sono di fatto rappresentate nella SI. Una spiegazione di questa apparente incongruenza viene dal fatto che, nelle reti neurali di tutte le stazioni delle vie sensitive, è presente un processo di inibizione laterale: ogni neurone, cioè, è collegato a quelli circostanti tramite neuroni internunciali inibitori, per cui esercita su di essi un’azione inibitoria la cui intensità cresce col crescere della sua frequenza di scarica. Ne deriva che i segnali tattili che provengono dalle regioni circostanti un punto cutaneo stimolato, tendono ad essere silenziati da quelli che provengono dal luogo ove cade lo stimolo. Questo si traduce, a livello corticale, in una maggior differenza tra l’attivazione dei neuroni centrali e quelli periferici nell’area che riceve ogni segnale tattile, quindi in una “restrizione funzionale” della sua estensione che fa sì che le aree corticali attivate dalla stimolazione di due punti vicini possano risultare spazialmente separate. E’evidente che l’intervento dell’inibizione laterale porta ad una maggior finezza e precisione nella rappresentazione sensoriale, quindi consente una miglior discriminazione spaziale degli stimoli.
Non va, altresì, dimenticato che la sovrapposizione delle aree sensoriali corticali ha riflessi funzionali analoghi a quelli descritti riguardo la sovrapposizione dei campi recettivi
periferici, ossia che possa aumentare la capacità di “riconoscere” i diversi caratteri degli stimoli. L’informazione su ciascun carattere arriva, infatti, alla corteccia suddivisa su un gruppo di fibre afferenti utilizzate “in condivisione” anche per gli altri caratteri secondo un codice che tenga conto della distribuzione delle frequenze di scarica nelle fibre del gruppo (“pattern code”). L’interpretazione dei messaggi sensoriali così codificati, al fine di estrarne tutte le informazioni (intensità, localizzazione ed altri caratteri), è affidato alla rete neuronica della corteccia della SI [1].