~ 4 ~
Introduzione
Sistemi di Memoria
Le memorie quotidiane definiscono chi siamo, senza memoria non potremmo imparare dalle nostre esperienze e il senso dell’identità personale che tutti possediamo sarebbe perso (Squire & Kandel, 1999).
La memoria rappresenta quindi la facoltà di immagazzinare e di avere accesso alle conoscenze apprese relative al mondo circostante. Tutti gli animali anche quelli più semplici possiedono la capacità di apprendere dall’ambiente che li circonda, questa facoltà nell’uomo raggiunge la sua massima complessità; la memoria umana infatti non è una capacità cognitiva unitaria, ma piuttosto è composta da un'intera serie di sistemi interconnessi complessi con funzioni differenti e specifiche, che sono supportati da substrati neurali anatomo-funzionali distinti. I risultati delle ricerche neuropsicologiche e neurobiologiche hanno portato la proposta dell'esistenza di sistemi di memoria multipli e dissociabili. Un sistema di memoria è definito come l’insieme dei processi portati avanti in aree cerebrali specifiche che permettono di immagazzinare o richiamare uno specifico tipo di informazione mnestica, attraverso un’attività separabile da quella di un diverso sistema (Berardi & Pizzorusso, 2006). Le principali evidenze sull'esistenza di sistemi di memoria separati, sono tratte da studi neuropsicologici, in cui era stata osservata una precisa corrispondenza tra specifici deficit di memoria e specifici pattern di lesione cerebrale. In questo modo è stato dimostrato che forme di memorie diverse sono dipendenti dall'integrità di strutture cerebrali specifiche che possono essere selettivamente danneggiate (Gaffan, 1974) (Squire, 1992).
A questi studi classici sono stati affiancati dati ottenuti con tecniche di neuro immagine che hanno permesso la visualizzazione dell’attività cerebrale in soggetti normali durante lo svolgimento di diversi compiti di memoria.
~ 5 ~
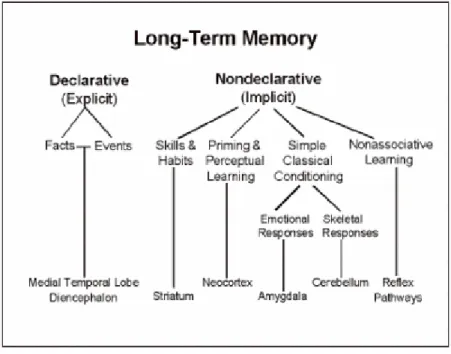
Sulla base di tali studi sono stati distinti dei grandi sistemi di memoria associati a substrati neurali differenti (figura 1-1): due di memoria a lungo termine, il sistema della memoria dichiarativa (o esplicita) e quello della memoria non dichiarativa (o implicita), e uno a breve termine cioè il sistema della memoria di lavoro. La memoria dichiarativa è l’insieme delle conoscenze a cui abbiamo accesso consapevole e si contrappone alla memoria non dichiarativa che invece raggruppa tutte quelle conoscenze alle quali non abbiamo accesso coscientemente, come le abilità motorie e percettive (memoria procedurale), i comportamenti appresi attraverso processi di condizionamento, e il priming.
Figura 0-1 Classificazione dei sistemi di memoria e delle principali strutture anatomiche che le sottendono (Squire, 1999)
La memoria dichiarativa può essere ulteriormente suddivisa in memoria riguardante gli eventi della nostra vita nel contesto spaziale e temporale in cui si sono verificati (memoria episodica) e in memoria riguardante le conoscenze relative al mondo estratte dal contesto (memoria semantica), le quali contribuiscono alla formazione di concetti, categorie e significati, compresa la conoscenza di noi stessi (Tulving, 1985). Una particolare forma di memoria dichiarativa è la memoria spaziale cioè un tipo di memoria che coinvolge abilità come il memorizzare mappe, punti di riferimento, e sviluppare rappresentazioni
~ 6 ~
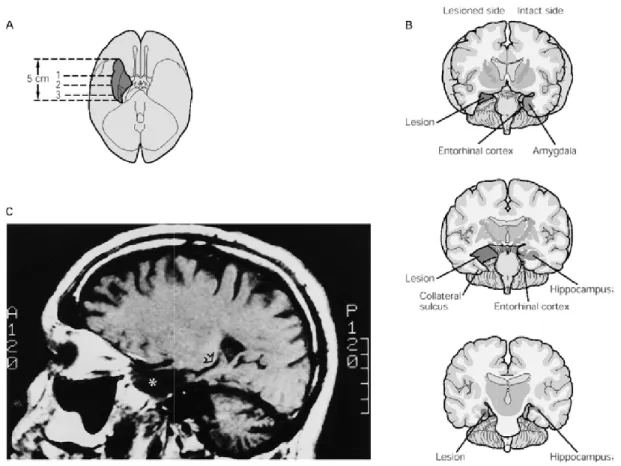
mentali dell’ambiente esterno , sebbene non debba essere dichiarata, le memorie topografiche possono essere dimostrate muovendosi con sicurezza da un posto all’atro all’interno di un ambiente conosciuto. I primi studi sulle memorie dichiarative furono fatti da Scoville e Milner (1957), che indagarono sugli effetti della rimozione bilaterale dell'ippocampo e delle vicine regioni del lobo temporale eseguita a scopo terapeutico in alcuni pazienti affetti da epilessia del lobo temporale. Il primo e più celebre caso fu quello del paziente H.M., che soffriva da oltre 10 anni di accessi epilettici intrattabili. In particolare il suo intervento chirurgico di asportazione bilaterale dei lobi temporali mediali interessò il complesso ippocampale, composto oltre che dalle regioni CA1, CA2, CA3, dal giro dentato e dal subiculum, anche dalla regione paraippocampale, dalla corteccia entorinale, peririnale e paraippocampale (Fig. 1-2). L'operazione risultò efficace nel controllo dell’epilessia, mantenne intatta la sua intelligenza, le sue funzioni percettive e motorie e le capacità mnestiche a breve termine, ma causò una grave amnesia anterograda, ovvero l'incapacità di formare nuove memorie, e una limitata amnesia retrograda, ovvero la perdita dei ricordi acquisiti nel periodo precedente al danno. Gli studi condotti sul paziente evidenziarono che questa incapacità di apprendere e memorizzare nuove informazioni era specifica per alcune forma di memoria che includevano le capacità di orientamento in spazi nuovi: H.M. mostrava di poter apprendere e mantenere in memoria alcune informazioni per breve tempo (ad esempio la capacità di ripetere dopo 30 secondi una breve lista di numeri memorizzati era normale) e di poter mantenere a lungo termine delle capacità apprese dopo l’intervento. Ad esempio, imparò ad eseguire dei movimenti fini con la mano osservandone i movimenti allo specchio e risultò anche in grado di migliorare con la pratica le prestazioni in molti altri compiti impliciti, ottenendo delle performance del tutto simili ai pazienti senza lesioni al lobo medio temporale, anche se, una volta interrogato sui compiti effettuati, non aveva memoria cosciente di averli eseguiti. Le osservazioni sperimentali indicavano quindi che nel paziente rimanevano intatte sia la memoria a breve termine che la memoria di lavoro che l’acquisizione di abilità manuali, con un ritmo di acquisizione e per un tempo di ritenzione paragonabile ai controlli sani.
~ 7 ~
Osservazioni successive hanno confermato l’amnesia anterograde vista in H.M., ed hanno individuato nell’ippocampo e nelle aree corticali circostanti le strutture del lobo medio temporale MTL cruciali per la formazione ed il consolidamento iniziale di nuove tracce di memoria esplicita a lungo termine (Scoville & Milner, 1957)
Si presume che delle due componenti della memoria esplicita, la memoria episodica, caratterizzata dalla presenza di forti elementi di contesto, sia mediata da un sistema composto dall’ippocampo e dalle sue connessioni con i corpi mammillari e con i nuclei del talamo anteriore, mentre la memoria semantica, priva di elementi di contesto, sembra essere supportata maggiormente dalla corteccia peririnale e dalle sue connessioni con il nucleo dorsomediale del talamo. Va sottolineato che la corteccia paraippocampale sembra essere particolarmente impegnata nella formazione di tracce di memoria spaziale (Rosenbaum, 2000). Dati più recenti hanno suggerito che non tutte le forme di memoria dichiarativa sono ugualmente compromesse dal danno ippocampale: Vargha-Khadem e collaboratori (1997) hanno riportato tre interessanti casi di amnesia anterograda che suggeriscono che le persone con una limitata lesione ippocampale possano acquisire memorie di tipo semantico (fatti, informazioni) e mantenere la memoria di familiarità sebbene mostrino gravi amnesie per quanto riguarda le informazioni episodiche. Lesioni totali dell’ippocampo e della corteccia limbica compromettono entrambi i tipi di memoria. Nonostante la critica agli studi di Vargha-Khadem che i pazienti da lei studiati, tutti in una fase precoce dello sviluppo al momento della lesione, potessero avere fenomeni di compensazione e che questo fosse il motivo della differenza con gli effetti della lesione al MTL nell’adulto, vi è adesso un consenso abbastanza generale sul fatto che la memoria semantica, inclusa la memoria di familiarità, data l’assenza di informazioni contestuali in tali tipi di memoria, possa sopravvivere a lesioni ippocampali, grazie ad una possibile dipendenza di tale forma di memoria da altre regioni del MTL (O'Kane & Kensinger, 2004) (Bayley & Squire, 2005).
Figura 0-2 Immagine del cervello del soggetto HM
L’intervento di HM provocò anche un
dichiarative, che interessava solo gli anni precedenti all manteneva intatti i ricordi d’infanzia.
casi di pazienti in cui, in seguito a una lesione d legata all’”età” di quest’
della memoria ed il momento della lesione
recenti è risultato superiore a quello sulle memorie remote (Rosenbaum et al., 2000) (Teng & Squire, 1999)
dallo psicologo francese, che suggerì che le memorie possono
organizzate attraverso il tempo affermando che in seguito a un danno temporale le nuove memorie vengono inficiate maggiormente rispetto a quelle acquisite precedentemente. Le conclusioni tratte da questi primi studi erano che le tracce memoria più remote non dipendevano più dall’ippocampo e dalle altre strutture del lobo temporale mediale, grazie alle quali si erano formate e consolidate, in
~ 8 ~
Immagine del cervello del soggetto HM (Kandel, 2003)
L’intervento di HM provocò anche un’amnesia retrograda parziale delle memorie che interessava solo gli anni precedenti all’intervento, mentre manteneva intatti i ricordi d’infanzia. In letteratura vengono riportati numerosi casi di pazienti in cui, in seguito a una lesione del MTL, la perdita di memoria è ’ultima, ovvero alla distanza temporale fra l’acquisizione della memoria ed il momento della lesione: generalmente l'effetto sui ricordi più recenti è risultato superiore a quello sulle memorie remote (Rosenbaum et al.,
(Teng & Squire, 1999). Questa dissociazione è nota come
dallo psicologo francese, che suggerì che le memorie possono essere gradualmente organizzate attraverso il tempo affermando che in seguito a un danno temporale le nuove memorie vengono inficiate maggiormente rispetto a quelle acquisite
Le conclusioni tratte da questi primi studi erano che le tracce memoria più remote non dipendevano più dall’ippocampo e dalle altre strutture del lobo temporale mediale, grazie alle quali si erano formate e consolidate, in
(Kandel, 2003)
amnesia retrograda parziale delle memorie intervento, mentre HM In letteratura vengono riportati numerosi MTL, la perdita di memoria è , ovvero alla distanza temporale fra l’acquisizione ralmente l'effetto sui ricordi più recenti è risultato superiore a quello sulle memorie remote (Rosenbaum et al., . Questa dissociazione è nota come gradiente di Ribot, essere gradualmente organizzate attraverso il tempo affermando che in seguito a un danno temporale le nuove memorie vengono inficiate maggiormente rispetto a quelle acquisite Le conclusioni tratte da questi primi studi erano che le tracce di memoria più remote non dipendevano più dall’ippocampo e dalle altre strutture del lobo temporale mediale, grazie alle quali si erano formate e consolidate, in
~ 9 ~
quanto erano state trasferite in altre strutture cerebrali. Come magazzino finale delle memorie dichiarative remote veniva proposta la corteccia frontale e le cortecce temporali. Tuttavia, con l’aumentare degli studi sull’argomento, questa semplice visione è stata messa in discussione. Esiste infatti una notevole variazione della gravità temporale dell’amnesia retrograda documentata in letteratura, la quale può spaziare da pochi mesi a diversi decenni (Rempel-Clower et al., 1996). Tale amnesia può dipendere da almeno due fattori in grado di spiegare questa variabilità. In primo luogo, l’entità della lesione: ad esempio, se il danno è limitato alla regione CA1 dell'ippocampo, l’amnesia retrograda potrà interessare gli ultimi 2 anni di vita. Al contrario, se la lesione è maggiormente estesa nel MTL, includendo l'ippocampo e parti della corteccia entorinale, l’amnesia potrà ampliarsi indietro nel tempo per un paio di decenni. In secondo luogo, questa gradualità potrebbe essere correlata al tipo di memoria dichiarativa testata, come dimostra la dissociazione tra i ricordi semantici ed episodici, i quali necessitano di un ippocampo intatto (Vargha-Khadem et al., 1997). Ci sono diverse difficoltà nello studio dell’amnesia retrograda in pazienti con danni al cervello, fondamentalmente perché questi studi si affidano prevalentemente a test retrospettivi le cui performance sono difficilmente comparabili nei diversi punti temporali; in più l’estensione del danno può variare da caso a caso e le lesioni sono raramente confinate ad una sola area (Squire & Alvarez, 1995). E’ per questi motivi che agli studi sull’uomo sono stati affiancati da studi su modelli animali, che presentano tra i principali vantaggi la possibilità di seguire gli studi sulle amnesie in maniera prospettica, controllare l’estensione della lesione e di conoscere l’origine e il tipo di memoria studiata. In questi studi sono stati utilizzati diversi modelli comportamentali per saggiare differenti sistemi mnestici, come il Morris water maze ( modello di memoria spaziale ippocampo dipendente), il fear Conditioning e il socially-acquired food preference (forme di memorie non spaziale). Sebbene questi protocolli comportamentali presentino delle profonde differenze in termini di proprietà dello stimolo, di motivazione e di richieste della performance, rappresentano tutti aspetti diversi della memoria dichiarativa umana, tutti infatti richiedono una complessa relazione tra gli stimoli e le
~ 10 ~
informazioni spaziali e temporali del contesto. Anche questi studi hanno mostrato in casi di lesione ippocampale (farmacologica o genetica) un impatto maggiore sulle memorie di recente formazione rispetto alle memorie remote.
Tuttavia, sulla base di evidenze più recenti, attualmente è in corso un acceso dibattito sulle strutture implicate nell’immagazzinamento e nel recupero di memorie esplicite remote, con possibili divergenze all’interno della stessa memoria episodica, in particolare sia per quanto riguarda gli aspetti spaziali e non (Kumaran & Maguire, 2005) che per i diversi aspetti della memoria spaziale (Kessel et al. 2001) (Rosenbaum, 2004) (Lopez, Herbeaux, & Bontempi, Context-Dependent Modulation of Hippocampal and Cortical Recruitment During Remote Spatial Memory Retrieval, 2011). Secondo alcuni studiosi, il recupero, di memorie dichiarative anche da molto tempo consolidate ma ricche di elementi di contesto, non si affrancherebbe mai dall’ippocampo (teoria della traccia multipla) e, in soggetti con lesioni ippocampali, la qualità del recupero delle memorie più antiche, pur presente, non sarebbe equivalente a quella di soggetti normali; secondo altri, invece, le memorie più antiche diventerebbero completamente indipendenti dall’ippocampo.
Nella parte seguente della tesi riassumerò brevemente le evidenze in letteratura relative alla formazione iniziale ed al consolidamento delle tracce di memorie dichiarative.
~ 11 ~
Formazione delle memorie
Le prime evidenze sull’esistenza di una rappresentazione anatomica dei processi mnemonici nell’uomo furono messe in luce dagli studi di Wilder Penfield, il quale osservò, attraverso la stimolazione elettrica della superficie cerebrale, che agendo sui lobi temporali otteneva quella che chiamava una risposta esperienziale, o esperienza retrospettiva, la quale consisteva nella descrizione da parte del paziente di un ricordo coerente di un'esperienza precedente. Il ruolo di tali zone venne ulteriormente chiarito, come già menzionato, all'inizio degli anni cinquanta dalle ricerche della sua collaboratrice, Brenda Milner, grazie agli effetti della rimozione bilaterale del lobo temporale mediale.
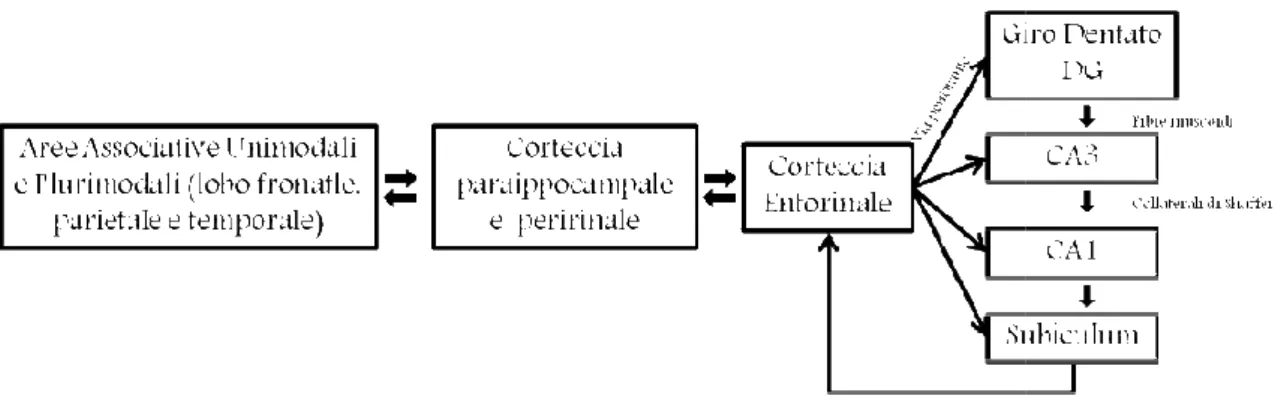
Ad oggi, è noto che l'informazione del mondo esterno arriva al nostro cervello tramite sistemi sensoriali che trasmettono i segnali alla neocorteccia, dove si formano le rappresentazioni sensoriali degli oggetti e degli eventi, questi segnali vengono inviati all’ippocampo. Il principale input alla formazione ippocampale proviene dalla corteccia entorinale i cui assoni terminano nel giro dentato e nei campi CA1 e CA3. La corteccia entorinale a sua volta riceve input dalle varie regioni associative neocorticali e dalla corteccia limbica sia direttamente che indirettamente tramite due regioni note come corteccia paraippocampale e peririnale, le quali integrano le informazioni provenienti dalle diverse modalità sensoriali e le inviano all'ippocampo propriamente detto; collettivamente queste tre regioni costituiscono la corteccia limbica del lobo temporale mediale (fig 1-3).
Figura 0-3 Rappresentazione grafica del “sistema delle memorie del lobo temporale
All'interno dell'ippocampo possono essere descritte tre vie connettive: il pathway delle via perforante, che proietta dalla corteccia entorinale alle cellule dei granuli del giro dentato; il pathway delle fibre muscoidi che contiene gli assoni delle cellule dei granuli che arrivano alle cellule piramidali della CA3 dell’ippocampo; e il pattern delle collaterali di Schaffer che consistono in collaterali eccitatorie delle cellule piramidali della CA3 che terminano sulle cellule piramidali della CA1 (Kandel, Principi di Neuroscienze, 2003)
sostenere fenomeni di potenziamento a lungo t
loro insieme costituiscono un circuito molto complesso, che partecipa all'elaborazione dei segnali in arrivo, detto circu
importante nella trasmissione di segnali dalla e per la corteccia entorinale (fig 1
~ 12 ~
Rappresentazione grafica del “sistema delle memorie del lobo temporale Principi di Neuroscienze, 2003)
dell'ippocampo possono essere descritte tre vie connettive: il pathway delle via perforante, che proietta dalla corteccia entorinale alle cellule dei granuli del giro dentato; il pathway delle fibre muscoidi che contiene gli assoni delle li che arrivano alle cellule piramidali della CA3 dell’ippocampo; e il pattern delle collaterali di Schaffer che consistono in collaterali eccitatorie delle cellule piramidali della CA3 che terminano sulle cellule piramidali della CA1
Neuroscienze, 2003). Tutte queste vie sono in grado di di potenziamento a lungo termine (Box 1). Questi pathway nel loro insieme costituiscono un circuito molto complesso, che partecipa all'elaborazione dei segnali in arrivo, detto circuito trisinaptico, particolarmente importante nella trasmissione di segnali dalla e per la corteccia entorinale (fig 1
Rappresentazione grafica del “sistema delle memorie del lobo temporale. (Kandel,
dell'ippocampo possono essere descritte tre vie connettive: il pathway delle via perforante, che proietta dalla corteccia entorinale alle cellule dei granuli del giro dentato; il pathway delle fibre muscoidi che contiene gli assoni delle li che arrivano alle cellule piramidali della CA3 dell’ippocampo; e il pattern delle collaterali di Schaffer che consistono in collaterali eccitatorie delle cellule piramidali della CA3 che terminano sulle cellule piramidali della CA1 Tutte queste vie sono in grado di Questi pathway nel loro insieme costituiscono un circuito molto complesso, che partecipa ito trisinaptico, particolarmente importante nella trasmissione di segnali dalla e per la corteccia entorinale (fig 1-4).
Figura 0-4 Tale schema è valido sia per i primati che per i roditori. L’ippocampo
informazioni dalla maggior parte delle aree associative neocorticali. Tali informazioni giungono attraverso le altre strutture di MTL, la corteccia peririnale e la corteccia paraippocampale, che a loro volta proiettano alla corteccia entorinale. Que
ippocampali, che le elaborano e le trasmettono, tramite la corteccia entorinale, alle aree neocorticali.
Gli output del sistema ippocampale hanno principalmente 2 origini proviene dagli assoni del
termina nell'adiacente campo CA1 e l'altro viaggia attraverso il fornice fino a raggiungere le strutture del proencefalo basale, inclusi il setto e i corpi mammilari. La seconda uscita proviene dagli assoni
subiculum che a sua volta proietta al di fuori della formazione ippocampale fino alla corteccia entorinale e anche
Un ulteriore input alla formazione ippocampale proviene da
sottocorticali attraverso il fornice, ma questo circuito dovrebbe avere solamente una funzione modulatoria.
Come si può dedurre da quanto detto le connessioni tra l'ippocampo neocorteccia sono tutte più o meno reciproche. Di conseguenza, l
trasportano le informazioni della neocorteccia alle aree all'ippocampo sono rispecchiate da vie che
della corteccia entorinale e terminano nelle stesse aree neocorticali che hanno originato gli input.
~ 13 ~
Tale schema è valido sia per i primati che per i roditori. L’ippocampo informazioni dalla maggior parte delle aree associative neocorticali. Tali informazioni giungono attraverso le altre strutture di MTL, la corteccia peririnale e la corteccia paraippocampale, che a loro volta proiettano alla corteccia entorinale. Questa invia le informazioni ai diversi circuiti ippocampali, che le elaborano e le trasmettono, tramite la corteccia entorinale, alle aree neocorticali.
Gli output del sistema ippocampale hanno principalmente 2 origini del campo CA3, che hanno due ramificazioni;
termina nell'adiacente campo CA1 e l'altro viaggia attraverso il fornice fino a raggiungere le strutture del proencefalo basale, inclusi il setto e i corpi mammilari.
proviene dagli assoni del campo CA1 che proiettano volta proietta al di fuori della formazione ippocampale fino alla corteccia entorinale e anche, attraverso la fimbria, fino al proencefalo basale.
put alla formazione ippocampale proviene da
sottocorticali attraverso il fornice, ma questo circuito dovrebbe avere solamente una funzione modulatoria.
dedurre da quanto detto le connessioni tra l'ippocampo neocorteccia sono tutte più o meno reciproche. Di conseguenza, l
trasportano le informazioni della neocorteccia alle aree del
all'ippocampo sono rispecchiate da vie che escono dall'ippocampo in direzione della corteccia entorinale e terminano nelle stesse aree neocorticali che hanno
Tale schema è valido sia per i primati che per i roditori. L’ippocampo riceve informazioni dalla maggior parte delle aree associative neocorticali. Tali informazioni giungono attraverso le altre strutture di MTL, la corteccia peririnale e la corteccia paraippocampale, che a sta invia le informazioni ai diversi circuiti ippocampali, che le elaborano e le trasmettono, tramite la corteccia entorinale, alle aree neocorticali.
Gli output del sistema ippocampale hanno principalmente 2 origini: la prima , che hanno due ramificazioni; un ramo termina nell'adiacente campo CA1 e l'altro viaggia attraverso il fornice fino a raggiungere le strutture del proencefalo basale, inclusi il setto e i corpi mammilari. CA1 che proiettano al volta proietta al di fuori della formazione ippocampale fino fino al proencefalo basale. put alla formazione ippocampale proviene dalle regioni sottocorticali attraverso il fornice, ma questo circuito dovrebbe avere solamente
dedurre da quanto detto le connessioni tra l'ippocampo e la neocorteccia sono tutte più o meno reciproche. Di conseguenza, le vie nervose che del MTL e quindi dall'ippocampo in direzione della corteccia entorinale e terminano nelle stesse aree neocorticali che hanno
~ 14 ~
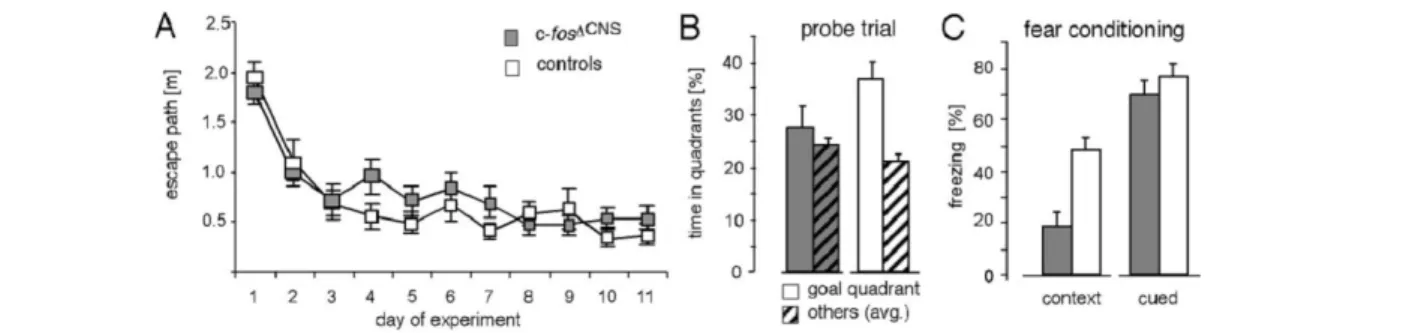
Box 1: Ruolo del potenziamento a lungo termineLo studio del potenziamento a lungo termine nella formazione ippocampale ha suggerito un meccanismo che potrebbe essere responsabile dei cambiamenti sinaptici che si verificano durante l'apprendimento e quindi potrebbe essere alla base della formazione delle memorie a lungo termine. Nel 1973 Lomo e Bliss scoprirono che uno stimolo ad alta frequenza in una qualsiasi delle 3 vie ippocampali dava origine a fenomeni di potenziamento a lungo termine dell’efficacia sinaptica (sia in vivo che in vitro) ma con caratteristiche differenti. Il potenziamento a lungo termine (LTP) nelle fibre muscoidi ha un carattere non associativo, dipende dall’ingresso di ioni calcio, e a seguito della stimolazione tetanica attiva la via dell’AMPc. Nella via Perforante e nelle collaterali di Shaffer c’è un LTP di tipo associativo, ovvero coinvolge la contemporanea attivazione del terminale postsinaptico e presinaptico, quindi dipende dalle peculiari caratteristiche dei recettori NMDA che controllano l’ingresso di ioni calcio che a sua volta porta all’attivazione di molecole enzimatiche dipendenti dal calcio come PKA, ERK, CaMKII. L’LTP è diviso in fasi successive, nella fase precoce che dura1-3 ore il calcio attiva molecole come la CaM-KII che è in grado di auto fosforilarsi e di indurre cambiamenti nell’attività, nel numero e nella localizzazione dei recettori AMPA per il glutammato, incrementando l’attività elettrica della sinapsi. Nella fase tardiva è richiesta la sintesi proteica realizzata mediante l’attivazione del pathway di CREB operata dalla stessa CaM-KII, la transizione dalla fase precoce alla fase tardiva è consentita dalla attivazione di molecole come PKA che fosforilano le fosfatasi inattivandole, in questo modo viene garantita l’attività degli attivatori di CREB. CREB induce a sua volta la trascrizione di molecole modulatorie e effettrici che promuovono meccanismi di plasticità come la formazione di nuove sinapsi. Sono stati utilizzati approcci genetici per indagare il ruolo di questi meccanismi sulla memoria. Un esempio rappresentativo viene da un lavoro di Fleischmann, et al. (2003) in cui sono stati utilizzati dei topi KO condizionali per il gene c-fos, un gene precoce rapidamente trascritto in seguito a fenomeni di plasticità attività dipendenti, la cui delezione era stata limitata alle regioni cerebrali, in primo luogo è stata osservata una riduzione della magnitudine dell’LTP tra la CA3 e la CA1 a causa di una riduzione nell’espressione del recettore NMDA, questo decremento è stato ritenuto responsabile della compromissione delle performance di richiamo della memoria in due test di apprendimento ippocampo dipendente, come il labirinto d’acqua di Morris (MWM) e il fear conditioning(FC). Questo studio, così come molti altri simili evidenziano che una riduzione della funzionalità di queste molecole è associato a forti deficit di formazione e mantenimento della traccia mnestica, collegati ad un significativo peggioramento delle performance in diversi compiti ippocampo dipendenti.
Figura b-1: Grafici delle perfomance dei topi c-fos∆SNC. A Le curve di apprendimento sono uguali tra i
KO e i WT. B e C si osserva un peggioramento delle performance nei KO rispetto ai controlli nel richiamo a 12 giorni sia nel MWM che nel FC (Fleischmann, et al., 2003).
~ 15 ~
In questo modo, le aree corticali interessate nell'elaborazione di uno stimolo possono, come vedremo in seguito, partecipare anche all’immagazzinamento a lungo termine delle memorie su quello stimolo. È importante sottolineare che grazie alle loro connessioni, le regioni del MTL fungono da zone di convergenza, che integrano le informazioni in maniera trasversale alle varie modalità sensoriali, creando così rappresentazioni che sono indipendenti dalla modalità originaria di elaborazione dell'informazione. Le regioni di convergenza sono implicate anche nella rappresentazione mentale, la quale permette di andare oltre la percezione fino ad avere una conoscenza formata da concetti e rappresentazioni astratte indipendenti dallo stimolo concreto, l'ippocampo infine riceve input da varie zone di convergenza delle regioni del MTL, elabora le informazioni ricevute e attraverso le sue connessioni efferenti modifica le memorie che si stanno consolidando, legandole insieme in modi che ci permettono di ricordare le relazioni tra gli elementi che le costituiscono anche se appresi in momenti differenti (Carlson, 2008). Senza dubbio l'ippocampo ha un ruolo essenziale nella
nostra capacità mnestica, tuttavia presenta dei limiti temporali
nell’immagazinamento e nel richiamo di certe forme di memorie, questa idea costituisce il punto centrale delle teorie classiche sul consolidamento che prevedono che ad una funzione ippocampale limitata debbano subentrare delle aree nuove che consentano lo stoccaggio a lungo termine per tali memorie. E abbastanza intuitivo immaginare l’esistenza di una migrazione della traccia dai circuiti ippocampali verso le aree di destinazione neocorticali, questa migrazione è quindi alla base del processo che consente l’assimilazione dell'esperienza fino alla sua memorizzazione permanente come un ricordo relativamente immune da perturbazioni (Lechner & Squire, 1999).
~ 16 ~
Il Consolidamento
Tutti i ricordi delle nostre esperienze di vita non si formano istantaneamente, le nuove memorie infatti sono gradualmente trasformate da uno stato inizialmente labile e prono all’estinzione ad uno stato più solido e permanente. Nel 1900 Müller e Pilzecker adottarono il termine “consolidamento” per descrivere il processo di trasformazione delle esperienze apprese in ricordi permanenti. Il consolidamento è un processo organizzato in due fasi successive, la prima più rapida che avviene a livello sinaptico, la seconda più lenta che coinvolge una riorganizzazione a livello sistemico (Dudai, 2004). Il cambiamento a livello sinaptico si avvia poche ore dopo l’apprendimento e include la crescita di nuove connessioni sinaptiche e il rimodellamento di connessioni preesistenti, così come avviene nei pathway ippocampali descritti precedentemente. Qualsiasi manipolazione sia genetica, farmacologica o ambientale che interferisce a qualunque livello di questo processo blocca la formazione della memoria a lungo termine, provocando la perdita della traccia di memoria entro poche ore dalla sua acquisizione (Kandel, 1998). Il consolidamento, però, per poter avvenire in maniera completa, oltre ad una riorganizzazione locale necessita, per quanto detto precedentemente, anche di una graduale riorganizzazione di intere regioni cerebrali che supportino le memorie dichiarative remote. Questa riorganizzazione avviene mediante il trasferimento del flusso di informazioni apprese attraverso la rete ippocampo-corticale. Durante alcuni studi di riproduzione simulata dell’attività mnestica, è stato visto che l’acquisizione di nuove informazioni era facilitata da una graduale acquisizione degli input, che consentiva di non creare delle interferenze tra le nuove e le vecchie memorie, implicando l’esistenza di una sede elaborazione primaria, rapida e sequenziale, in grado di trasferire le informazioni ad una sede secondaria di stoccaggio (McLelland & McNaughton, 1995). Questo processo di apprendimento usufruisce di una rete, che attraverso molte ripetizioni, definisce una nuova rappresentazione che va ad arricchire le conoscenze di base. Nei sistemi fisiologici è l’ippocampo a svolgere la codifica iniziale e a dirigere il processo di immagazzinamento corticale, partecipando, quando le esperienze
~ 17 ~
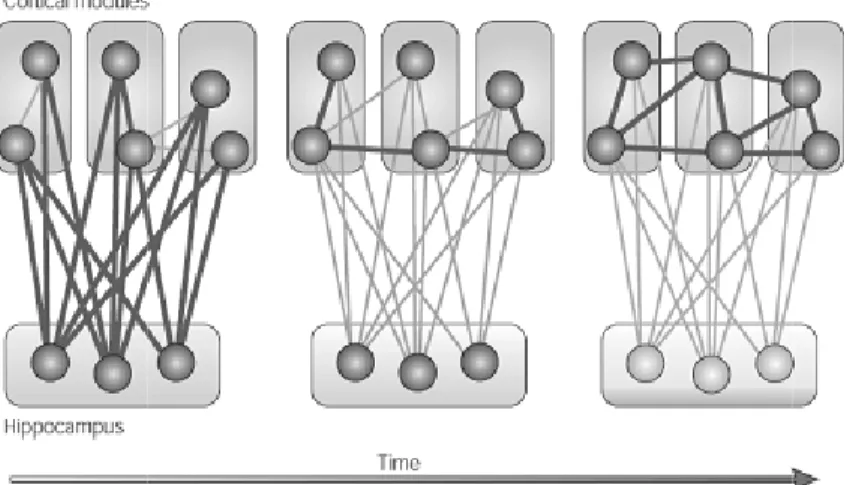
acquisite si ripropongono, al ripristino dei pattern di attivazione corticali precedentemente formati, ogni ripristino cambia le sinapsi corticali, in modo che l' acquisizione di una nuova conoscenza non interferisca con i ricordi precedenti e si consolidi. L’insieme degli studi sulle amnesie umane e sui modelli animali hanno consentito la formulazione di diversi modelli di consolidamento. Marr (1970) è stato il primo a proporre che l'ippocampo processi rapidamente gli eventi giornalieri prima che le informazioni vengano trasferite alla corteccia per la riorganizzazione e la successiva riclassificazione; inoltre ha proposto che il processo di trasferimento dipenda da un replay durante il sonno dell’attività neuronale corrispondente ai pattern memorizzati durante la veglia. Tali idee costituiscono la base dei modelli contemporanei di consolidamento della memoria. Secondo questi modelli cosiddetti classici o standard, l’ippocampo integra le informazioni di distribuiti moduli corticali, che rappresentano i vari caratteri dell’esperienza, per convertirli in una traccia di memoria coerente: tali cambiamenti sono rapidi e transienti. Una successiva riattivazione della rete ippocampo-corteccia porta ad un graduale rafforzamento delle connessioni cortico-corticali e alla creazione di nuove. Il rafforzamento incrementale di queste connessioni permette ai nuovi ricordi di diventare indipendenti dall’ippocampo e di essere progressivamente integrati con le memorie pre-esistenti. Queste memorie corticali diventano più resistenti al tempo e alle interferenze rispetto a quelle ippocampali. La caratteristica fondamentale di questo modello è che i cambiamenti della forza tra le connessioni del sistema ippocampale e delle diverse aree corticali sono rapidi e transitori, mentre le modifiche delle connessioni all’interno delle aree corticali sono lente e resistenti nel tempo.
Figura 0-5 Modello Standard di Consolidamento (Bontempi et al., 2005)
Sebbene non possa essere messa in discussione la funzione precoce dell’ippocampo questo mod
contradette da evidenze sperimentali successive.
La prima asserzione dice che una volta completato il consolidam connessioni ippocampali
elaborate e essendo transienti vengono rapidamente cancellate per consentirne il riciclo nella codifica di nuove memorie
asserzione, se non può essere considerata errata, quantomeno è considerabile come eccessivamente semplificata. Per esempio se da un lato sembra essere supportato dalle evidenze sulle amnesie retrograde dopo i danneggiamenti ippocampali i range di memorie perdute fatte risalire tra i 10 e i 20 anni vanno oltre la comune definizione d
La seconda asserzione del modello non prevede che ci sia alcuna differenza tra i meccanismi delle memorie semantiche e quelli delle memorie episodiche, e quindi che l’ippocampo abbia un ruolo importante
consolidamento non abbia alcun ruolo in entrambe esserci un’inesattezza come dimostrano gli studi di
precedentemente. Inoltre i risultati ottenuti da studi, in cui vengono recuperati ricordi remoti in soggetti con danno ippocampale, sia su umani (Rosenbaum et al., 2000) che su animali (Bontempi & Laurernt
~ 18 ~
Modello Standard di Consolidamento (Bontempi et al., 2005)
Sebbene non possa essere messa in discussione la funzione precoce dell’ippocampo questo modello prevede una serie implicazioni che sono state
da evidenze sperimentali successive.
La prima asserzione dice che una volta completato il consolidam
connessioni ippocampali, non sono più necessarie per il richiamo delle memorie elaborate e essendo transienti vengono rapidamente cancellate per consentirne il riciclo nella codifica di nuove memorie. I nuovi dati suggeriscono che questa asserzione, se non può essere considerata errata, quantomeno è considerabile ome eccessivamente semplificata. Per esempio se da un lato sembra essere supportato dalle evidenze sulle amnesie retrograde dopo i danneggiamenti ippocampali i range di memorie perdute fatte risalire tra i 10 e i 20 anni vanno oltre la comune definizione di “meccanismo transiente”.
a seconda asserzione del modello non prevede che ci sia alcuna differenza tra i meccanismi delle memorie semantiche e quelli delle memorie episodiche, e quindi
un ruolo importante nella codifica iniziale e
on abbia alcun ruolo in entrambe, anche in questo caso sembra esserci un’inesattezza come dimostrano gli studi di Vargha-Khandem
Inoltre i risultati ottenuti da studi, in cui vengono recuperati ricordi remoti in soggetti con danno ippocampale, sia su umani (Rosenbaum et al., (Bontempi & Laurernt-Demirc., 1999), indicano che la qualità
Modello Standard di Consolidamento (Bontempi et al., 2005)
Sebbene non possa essere messa in discussione la funzione precoce icazioni che sono state
La prima asserzione dice che una volta completato il consolidamento le iamo delle memorie elaborate e essendo transienti vengono rapidamente cancellate per consentirne il I nuovi dati suggeriscono che questa asserzione, se non può essere considerata errata, quantomeno è considerabile ome eccessivamente semplificata. Per esempio se da un lato sembra essere supportato dalle evidenze sulle amnesie retrograde dopo i danneggiamenti ippocampali i range di memorie perdute fatte risalire tra i 10 e i 20 anni vanno
a seconda asserzione del modello non prevede che ci sia alcuna differenza tra i meccanismi delle memorie semantiche e quelli delle memorie episodiche, e quindi ziale e che dopo il , anche in questo caso sembra Khandem illustrati Inoltre i risultati ottenuti da studi, in cui vengono recuperati ricordi remoti in soggetti con danno ippocampale, sia su umani (Rosenbaum et al., , indicano che la qualità
~ 19 ~
del ricordo non è la stessa presente in soggetti sani che invece riflettono il contributo importante del sistema ippocampale (Nadel & Booth, 2001) .
In risposta alle inesattezze del modello standard di consolidamento Nadel, Moscovitch e collaboratori proposero un modello alternativo che prese il nome di multiple trace theory, MTT. Questo modello sosteneva che l’ippocampo resta sempre implicato nell’immagazzinamento mnestico e nel richiamo di una rappresentazione contestuale e spaziale ricca di dettagli a prescindere dall’età della memoria. In particolare il complesso ippocampale codifica rapidamente tutte le informazioni per poi andare a creare una traccia di memoria sostenuta dal legame con i neuroni neocorticali: di conseguenza la memoria di un episodio consisterà in un network ippocampo-corteccia. Ogni volta che la memoria precedentemente immagazzinata è recuperata, con il coinvolgimento di entrambi i siti, ippocampali e neocorticali, viene attivata una nuova codifica all’interno dell'ippocampo. Questa riattivazione della memoria genera tracce multiple nuove nell'ippocampo che proliferano in maniera distribuita, coinvolgendo sempre più regioni cerebrali, specialmente aree neocorticali, in modo da rendere i vecchi ricordi sempre più resistenti al decadimento o a seguito di un danno al sistema ippocampale (Nadel & Booth, 2001).
A sostegno di questo modello ci sono moltissimi studi di fMRI che mettono in evidenza l’attivazione funzionale dell’ippocampo in risposta a ricordi episodici fortemente legati al contesto sia di natura recente che remota, rispetto a ricordi di natura semantica. L’analisi è stata compiuta su un gruppo misto tra i 65 e i 70 anni precedentemente sottoposti ad un questionario che potesse fornire informazioni su esperienze personali nel tempo. Ai pazienti veniva richiesto di immaginare eventi correlati ad un cue che potesse riportare ad eventi personali. I risultati della risonanza hanno evidenziato un’ uguale attivazione dell’intera area ippocampale sia in risposta ad un richiamo di memorie recenti che a un richiamo di memorie remote di eventi autobiografici (fig 1-6).
Figura 0-6 A sinistra il grafico mostra l’attivazione dell’ippocampo durante il richiamo di memorie recenti e
remote richiamate da cue ricollegabili ad eventi personali a confronto con cue generici. A destra imagine della risonanza magnetic funzionale
Secondo la MTT il complesso ippocampale è quindi richiesto nel richiamo delle memorie episodiche per due ragioni fondamentali, in primo luogo perché gli episodi sono definiti dal contesto spaziale e temporale e il contesto è immagazzinato nell’ippocampo, e secondariamente perché sono le tracce del contesto a catalogare i dettagli contenuti nelle memorie episodiche, come si evince dagli studi che descriveremo in seguito (Ryan et al., 2001). Per distinguere più nettamente i ricordi legati al contes
studio analogo che utilizzava la fMRI, in cui in seguito alla visione di un video veniva richiesto di individuare degli oggetti presenti nel filmato, posizionare nello spazio degli elementi ed riordinare p
mostravano che l’ippocampo era preferenzialmente attivato durante il richiamo di informazioni contestuali spaziali.
importante in tutti i tipi di memoria dichiarativa ep
particolare importanza nelle memorie spaziali. Questo ruolo privilegiato fu messo in luce, da O’Keefe e Dostrovky nel 1971 con le prime evidenze sull’esistenza delle “place cell”, par
internamente delle rappresentazioni spaziali dell’ambiente circostante. Attualmente sappiamo che più che definire solamente una mappa spaziale l’ippocampo è dedicato ad identificare relazioni e nessi tra oggetti, fatti e eventi (Shreger et al., 2007). Più specificamente i
~ 20 ~
A sinistra il grafico mostra l’attivazione dell’ippocampo durante il richiamo di memorie recenti e remote richiamate da cue ricollegabili ad eventi personali a confronto con cue generici. A destra imagine della
risonanza magnetic funzionale (Nadel & Booth, 2001).
Secondo la MTT il complesso ippocampale è quindi richiesto nel richiamo delle memorie episodiche per due ragioni fondamentali, in primo luogo perché gli episodi sono definiti dal contesto spaziale e temporale e il contesto è ppocampo, e secondariamente perché sono le tracce del contesto a catalogare i dettagli contenuti nelle memorie episodiche, come si evince dagli studi che descriveremo in seguito (Ryan et al., 2001). Per distinguere più nettamente i ricordi legati al contesto, spaziale e temporale, è stato effettuato uno studio analogo che utilizzava la fMRI, in cui in seguito alla visione di un video veniva richiesto di individuare degli oggetti presenti nel filmato, posizionare nello spazio degli elementi ed riordinare particolari eventi avvenuti nel video. I risult mostravano che l’ippocampo era preferenzialmente attivato durante il richiamo di informazioni contestuali spaziali. L’ippocampo ha chiaramente un ruolo importante in tutti i tipi di memoria dichiarativa episodica, riveste però una particolare importanza nelle memorie spaziali. Questo ruolo privilegiato fu messo in luce, da O’Keefe e Dostrovky nel 1971 con le prime evidenze sull’esistenza delle “place cell”, particolari neuroni ippocampali in grado di gene internamente delle rappresentazioni spaziali dell’ambiente circostante. Attualmente sappiamo che più che definire solamente una mappa spaziale l’ippocampo è dedicato ad identificare relazioni e nessi tra oggetti, fatti e eventi
Più specificamente i net-work ippocampali codif
A sinistra il grafico mostra l’attivazione dell’ippocampo durante il richiamo di memorie recenti e remote richiamate da cue ricollegabili ad eventi personali a confronto con cue generici. A destra imagine della
Secondo la MTT il complesso ippocampale è quindi richiesto nel richiamo delle memorie episodiche per due ragioni fondamentali, in primo luogo perché gli episodi sono definiti dal contesto spaziale e temporale e il contesto è ppocampo, e secondariamente perché sono le tracce del contesto a catalogare i dettagli contenuti nelle memorie episodiche, come si evince dagli studi che descriveremo in seguito (Ryan et al., 2001). Per distinguere più to, spaziale e temporale, è stato effettuato uno studio analogo che utilizzava la fMRI, in cui in seguito alla visione di un video veniva richiesto di individuare degli oggetti presenti nel filmato, posizionare nello
articolari eventi avvenuti nel video. I risultati mostravano che l’ippocampo era preferenzialmente attivato durante il richiamo di L’ippocampo ha chiaramente un ruolo isodica, riveste però una particolare importanza nelle memorie spaziali. Questo ruolo privilegiato fu messo in luce, da O’Keefe e Dostrovky nel 1971 con le prime evidenze sull’esistenza neuroni ippocampali in grado di generare internamente delle rappresentazioni spaziali dell’ambiente circostante. Attualmente sappiamo che più che definire solamente una mappa spaziale l’ippocampo è dedicato ad identificare relazioni e nessi tra oggetti, fatti e eventi work ippocampali codificano i
~ 21 ~
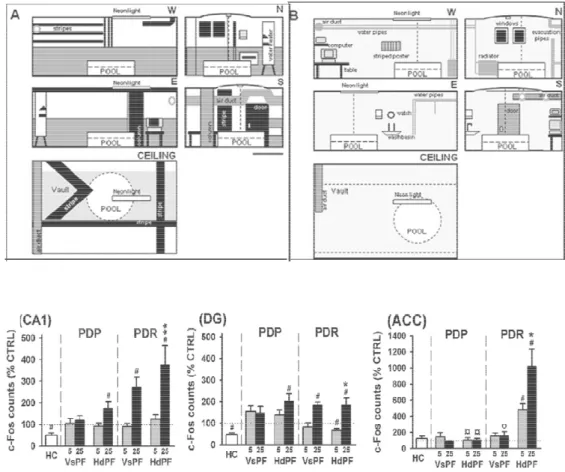
percorsi attraverso lo spazio così come fanno per gli episodi di memoria, i quali sono definiti come sequenze di eventi e di luoghi in cui questi episodi si verificano. Quindi lo studio del ruolo dell’ippocampo nella memoria spaziale, può essere visto come esempio specifico della capacità generale dell’ippocampo che comprende tutta la memoria dichiarativa ed in particolare, il richiamo di rappresentazioni molto dettagliate dell’ambiente (Shreger et al., 2007). Alcuni degli studi sugli animali, effettuati fin’ora, mostrano un quadro discordante, in questi vari studi gli animali sperimentali vengono addestrati a un compito spaziale ed in seguito la memoria recente viene tipicamente testata dopo un giorno dal training, mentre la memoria remota dopo 25-30 giorni. I metodi maggiormente usati sono il monitoraggio dell’attivazione metabolica a livello regionale, segno della riattivazione della traccia di memoria, il controllo dell’espressione genetica attività-regolata l’uso di modelli animali con anomalie nella plasticità corticale. I dati disponibili hanno riscontrato che il richiamo delle memorie spaziali recenti è stato associato con l'attivazione dell'ippocampo, mentre il richiamo di ricordi remoti è stato prevalentemente associato con l'attivazione di regioni corticali come la corteccia prefrontale, frontale, cingolata anteriore (Frankland et al., 2001) (Maviel & Bontempi, Sites of neocortical reorganization critical for Remote Spatial Memory, 2004).
Questi lavori, che vedremo successivamente nel dettaglio, forniscono prove a favore della teoria standard di consolidamento ed in particolare evidenzia il ruolo delle aree prefrontali nello stoccaggio e del richiamo delle memorie remote. Tuttavia lavori più recenti rimettono in discussione il decadimento ippocampale nel richiamo delle memorie remote, Bontempi e collaboratori (2011) hanno utilizzato un particolare protocollo del task comportamentale del Morris water maze in cui è stato saggiato il reclutamento dell’ippocampo e delle strutture prefrontali nel richiamo delle memorie remote in un compito d’apprendimento “difficile” (contesto distante e complesso) rispetto ad uno “facile”(contesto vicino e semplificato) comparando i livelli di attivazione di c-fos ippocampale durante il recupero di memorie remote nei due compiti (fig 1-7). Nel contesto difficile l’ippocampo esprime un aumento dell’espressione di c-fos nel richiamo remoto
rispetto a quello recente,
il cui contemporaneo ingaggio riflette il dialogo tra queste due regioni.
Figura 0-7 Le figura in alto mostra i due apparati utilizzati nel
Performance Degradetion Resistant (PDR) corrispondente ad un ambiente con contesto complesso; B corrisponde all’apparato definito come Performance Degradetion Prone (PDP) corrispondente ad un ambiente semplice. I grafici in basso riportano i risultati dell’attivazione di c
un’area prefrontale aCC. Per ciascun ambiente e in corrispondenza di due punti temporali, uno recente 5 giorni e uno remoto 25 giorni, sono stati utili
nascosta e l’altro con piattaforma visibile, entrambi razionalizzati sull’attivazione in un gruppo di controllo home cage. (Bontempi et al., 2011)
La divergenza tra questi risultati, e di conseguenza tra le due teorie, potrebbe essere dovuta alla particolare peculiarità funzionale dell’ippocampo nel trattamento delle informazioni ricche di elementi
al.,2006; Winocur et al.,2010)
mostrato che il richiamo di precise memorie remote associate alle paure non richiede l’ippocampo, similmente risultati di fMRI
navigazione semplice in soggetti sani ha
dell’ippocampo e l’attivazione di diverse aree corticali a seconda del
~ 22 ~
rispetto a quello recente, in modo simile all’attivazione nella corteccia prefrontale, il cui contemporaneo ingaggio riflette il dialogo tra queste due regioni.
Le figura in alto mostra i due apparati utilizzati nel training A corrisponde all’apparato definito Performance Degradetion Resistant (PDR) corrispondente ad un ambiente con contesto complesso; B corrisponde all’apparato definito come Performance Degradetion Prone (PDP) corrispondente ad un ambiente I grafici in basso riportano i risultati dell’attivazione di c-fos in due aree ippocampali CA1 e DG e in un’area prefrontale aCC. Per ciascun ambiente e in corrispondenza di due punti temporali, uno recente 5 giorni e uno remoto 25 giorni, sono stati utilizzati due gruppi sperimentale uno testato con piattaforma nascosta e l’altro con piattaforma visibile, entrambi razionalizzati sull’attivazione in un gruppo di controllo home cage. (Bontempi et al., 2011)
La divergenza tra questi risultati, e di conseguenza tra le due teorie, potrebbe essere dovuta alla particolare peculiarità funzionale dell’ippocampo nel trattamento delle informazioni ricche di elementi contestualizzanti
et al.,2010). Wang e collaboratori (2009) recentemente hanno mostrato che il richiamo di precise memorie remote associate alle paure non richiede l’ippocampo, similmente risultati di fMRI (fig 1-8) in un compito di navigazione semplice in soggetti sani hanno mostrato il disimpegno
attivazione di diverse aree corticali a seconda del
in modo simile all’attivazione nella corteccia prefrontale, il cui contemporaneo ingaggio riflette il dialogo tra queste due regioni.
training A corrisponde all’apparato definito Performance Degradetion Resistant (PDR) corrispondente ad un ambiente con contesto complesso; B corrisponde all’apparato definito come Performance Degradetion Prone (PDP) corrispondente ad un ambiente fos in due aree ippocampali CA1 e DG e in un’area prefrontale aCC. Per ciascun ambiente e in corrispondenza di due punti temporali, uno recente 5 zzati due gruppi sperimentale uno testato con piattaforma nascosta e l’altro con piattaforma visibile, entrambi razionalizzati sull’attivazione in un gruppo di controllo
La divergenza tra questi risultati, e di conseguenza tra le due teorie, potrebbe essere dovuta alla particolare peculiarità funzionale dell’ippocampo nel contestualizzanti (Moscovitch et recentemente hanno mostrato che il richiamo di precise memorie remote associate alle paure non in un compito di nno mostrato il disimpegno attivazione di diverse aree corticali a seconda del compito,
anche se la corteccia paraippocampale sembra essere fondamentale per qualsiasi rappresentazione topografica (Rosenbaum et al., 2004)
Figura 0-8 Immagini di fMRI che riportano l’attivazione media tra tutti i partecipanti delle regioni cerebrali
riportate in tutti i tasks di memoria remota spaziale. A destra il giro paraippocampale; al
retro spinale; a destra la corteccia occipitale superiore. L’ippocampo non è stato attivo in nessun tasks
E’ chiaro che le memorie non sono un sistema unitario, e similmente anche le stesse memorie episodiche e più in particolare le memorie spaziali non possono essere considerate come tali, di conseguenza sono caratterizzate da molteplici specializzazioni supportate da substrati differenti
conseguenza i diversi tasks,
limitano all’ippocampo e le teorie esistenti non riescono a cogliere appieno la complessità del meccanismo dell’immagazzinamento e del richiamo delle varie forme di memoria. Quello che sembr
che aree neocorticali differenti costituiscon
impegnati in modo cooperativo attraverso l’ippocampo a fornire percorsi complementari per lo stoccaggio e il recupero delle memori
~ 23 ~
anche se la corteccia paraippocampale sembra essere fondamentale per qualsiasi rappresentazione topografica (Rosenbaum et al., 2004).
Immagini di fMRI che riportano l’attivazione media tra tutti i partecipanti delle regioni cerebrali riportate in tutti i tasks di memoria remota spaziale. A destra il giro paraippocampale; al
retro spinale; a destra la corteccia occipitale superiore. L’ippocampo non è stato attivo in nessun tasks (cerchio) (Rosenbaum et al., 2004)
E’ chiaro che le memorie non sono un sistema unitario, e similmente anche le pisodiche e più in particolare le memorie spaziali non possono essere considerate come tali, di conseguenza sono caratterizzate da molteplici specializzazioni supportate da substrati differenti. Le diverse codifiche e di conseguenza i diversi tasks, si basano su diversi substrati neurali che non si limitano all’ippocampo e le teorie esistenti non riescono a cogliere appieno la complessità del meccanismo dell’immagazzinamento e del richiamo delle varie forme di memoria. Quello che sembra chiaro e comunemente accettato è il fatto che aree neocorticali differenti costituiscono dei sistemi multipli e interattivi impegnati in modo cooperativo attraverso l’ippocampo a fornire percorsi complementari per lo stoccaggio e il recupero delle memorie (Ryan et al., 2009). anche se la corteccia paraippocampale sembra essere fondamentale per qualsiasi
Immagini di fMRI che riportano l’attivazione media tra tutti i partecipanti delle regioni cerebrali riportate in tutti i tasks di memoria remota spaziale. A destra il giro paraippocampale; al centro, la corteccia
retro spinale; a destra la corteccia occipitale superiore. L’ippocampo non è stato attivo in nessun tasks
E’ chiaro che le memorie non sono un sistema unitario, e similmente anche le pisodiche e più in particolare le memorie spaziali non possono essere considerate come tali, di conseguenza sono caratterizzate da molteplici Le diverse codifiche e di si basano su diversi substrati neurali che non si limitano all’ippocampo e le teorie esistenti non riescono a cogliere appieno la complessità del meccanismo dell’immagazzinamento e del richiamo delle varie
a chiaro e comunemente accettato è il fatto istemi multipli e interattivi impegnati in modo cooperativo attraverso l’ippocampo a fornire percorsi
~ 24 ~
I siti delle memorie remote
La ricerca dei sistemi specifici coinvolti nel consolidamento della memoria è iniziata, come già detto, con la constatazione che danni al MTL producono una grave amnesia anterograda e retrograda, ovvero, i pazienti riportavano grande difficoltà nella formazione di nuovi ricordi e nel recupero di ricordi recenti ma non delle memorie molto antiche. Una seconda osservazione, complementare alla precedente, è che, quando la patologia cerebrale comprende danni anche alla neocorteccia, la memoria remota è spesso compromessa (Squire et al., 2001). Ciò suggerisce che le aree neocorticali fungono da siti di stoccaggio per le memorie remote. Come abbiamo già assodato studi di apprendimento e memoria hanno fornito molte prove sul coinvolgimento di meccanismi ippocampali nell’acquisizione, nel mantenimento e nel recupero di memorie episodiche, tuttavia, abbiamo dato pochi accenni su come e dove queste informazioni vengono memorizzate in modo permanente. Una recente serie di studi molecolari e cellulari su roditori, con l’uso di diversi paradigmi comportamentali, hanno fornito convincenti prove sul coinvolgimento di specifiche regioni neocorticali nella memorizzazione di informazioni inizialmente strettamente legate all'ippocampo (Bontempi et al. 1999; Frankland et al. 2001, 2004; Maviel et al. 2004). In particolare le aree maggiormente interessate nel recupero delle memorie remote spaziali sembrano essere quelle della corteccia prefrontale, compresa la corteccia cingolata anteriore. C’è da sottolineare che gli studi su animali da laboratorio sono riusciti a fare luce sui siti di stoccaggio della memoria remota spaziale, anche se molto rimane ancora da approfondire per quanto riguarda il decorso temporale delle varie interazioni ippocampo-neocorteccia che rendono possibile la memoria a lungo termine, come, ad esempio quali sono i punti temporali precisi in cui le varie aree neocorticali vengono ingaggiate. Dato che dagli studi di amnesia retrograda, osservata in seguito a lesioni dell'ippocampo, nell'uomo e negli animali, è possibile dedurre solamente il ruolo da parte dell’ippocampo nell’immagazzinamento della memoria, Bontempi e collaboratori (1999), usando il monitoraggio dell’attività metabolica, hanno esaminato in che modo i circuiti
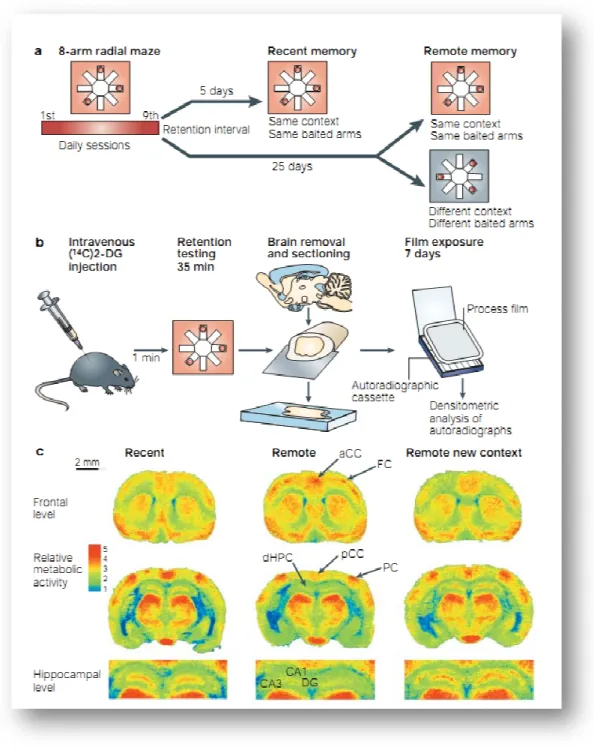
~ 25 ~
cerebrali sottostanti alla memoria a lungo termine sono riorganizzati nel tempo in un cervello intatto. Per fare questo hanno addestrato alcuni topi wild tipe ad un compito di apprendimento spaziale ippocampo-dipendente (eight-arms radial maze) ed hanno monitorato l’attività cerebrale durante il seguente recupero di una delle memorie o recenti, a 5 giorni dal training, o remote, a 25 giorni dal training. Il recupero delle memorie recenti spaziali ha prodotto una robusta attivazione dell'ippocampo rispetto a quello dei ricordi remoti, in cui viene persa la correlazione tra l’attività metabolica e la performance mnestica, risultato coerente con il modello che propone un progressivo disimpegno dell'ippocampo durante il consolidamento della memoria. Al contrario, diverse aree della neocorteccia, tra cui corteccia frontale, anteriore cingolata e temporale, hanno mostrato un pattern di attivazione opposto, queste aree non si attivavano in contesti differenti da quello appreso (Fig. 1-9). Questi risultati sono prove convincenti a favore della partecipazione di reti neocorticali nell’immagazzinamento delle memorie remote e di una riorganizzazione tempo-dipendente dei circuiti neuronali sottostanti alla memoria a lungo termine.
Figura 0-9 Riorganizzazione tempo
spaziali. A Protocollo comportamentale (gruppo di controllo testato in un ambiente differente).B Procedura di iniezione ((14C)2-DG) permette di visualizzare il cambiamento del
autoradiografico 148. C sezioni colorate co
l’attività è localizzata nelle aree ippocampali, di contro in corrispondenza delle memorie remote l’atti localizzata nelle aree frontali. (Bontempi et al.,1999).
~ 26 ~
tempo-dipendente dei circuiti neurali che supportano le memorie discriminative spaziali. A Protocollo comportamentale (gruppo di controllo testato in un ambiente differente).B Procedura di DG) permette di visualizzare il cambiamento dell’attività metabolica per mezzo del metodo autoradiografico 148. C sezioni colorate con metodo autoradiografico. I corrispondenza delle memorie recenti l’attività è localizzata nelle aree ippocampali, di contro in corrispondenza delle memorie remote l’atti
localizzata nelle aree frontali. (Bontempi et al.,1999).
che supportano le memorie discriminative spaziali. A Protocollo comportamentale (gruppo di controllo testato in un ambiente differente).B Procedura di l’attività metabolica per mezzo del metodo metodo autoradiografico. I corrispondenza delle memorie recenti l’attività è localizzata nelle aree ippocampali, di contro in corrispondenza delle memorie remote l’attivazione è
~ 27 ~
Sulla stessa linea adottata da Bontempi sono stati sviluppati moltissimi studi mirati ad individuare più specificamente le diverse aree coinvolte. In un lavoro del (2004) Maviel, combinando l’immaging funzionale dell’espressione genica con specifiche inattivazioni neuronali transitorie, in modo da ridurre al minimo eventuali meccanismi di compensazione associati a lesioni irreversibili, ha identificato la corteccia prefrontale e la corteccia anteriore cingolata come le aree critiche per la conservazione e il recupero di ricordi spaziali. Hanno mappato l’espressione di due geni precoci, Zif268 e c-fos che possono essere usati come marker dell’attivazione neuronale, molto utile per tracciare i cambiamenti a diversi punti temporali in seguito all’apprendimento, o come indice di un LTP in corso. I soggetti sono stati addestrati nel labirinti a 5 braccia ed è stato effettuato il richiamo della memoria a recente 1 giorno e remota 30 giorni. Lo studio ha interessato per la prima volta un ampio numero di aree, mai considerate precedentemente (fig. 1-10) mettendo in evidenza la partecipazione di un’intera rete di aree coinvolte in tempi differenti nei meccanismi di consolidamento. Si osserva una divergenza del coinvolgimento delle aree prefrontali e retrospleniali che si attivano in corrispondenza delle memorie remote opposte dalle aree posteriori, ippocampali ed entorinali che si attivano con le memorie recenti. Per confermare queste dissociazioni funzionali tra le regioni cerebrali, gli autori hanno anche utilizzato l’inattivazione farmacologica con lidocaina, bloccante dei canali sodio che sopprime l’attività neuronale inibitoria ed eccitatoria, in seguito all’addestramento di due nuovi gruppi di animali, ai quali selettivamente veniva danneggiata o un’area prefrontale o l’ippocampo (fig1-11).
Figura 0-10 Espressione di c-fos nella corteccia
cingolata posteriore, nella corteccia retrospleniale nell’ippocampo dorsale e ventrale, nel subiculum e nella corteccia entorinale dopo il test di memoria recente (giorno 1) o remoto (30 gi
microfotografie. L’espressione di c
che nella corteccia retrospinale, dove solo i dati di Zif268 rivelano un coinvolgimento di quest’area.(Maviel et al., 2004).
Figura 0-11 Effetti dell’inattivazione neuronale della corteccia prefrontale, cingolata anteriore, posteriore e
dell’ippocampo dorsale attraverso lidocaina (barre in nero) rispetto a un liquido
(aCSF, barre in bianco), nel test di memoria recente (giorno 1) o remota (giorno 30). (Maviel et al., 2004).
~ 28 ~
fos nella corteccia prefrontale, nella corteccia cingolata anteriore, nella corteccia cingolata posteriore, nella corteccia retrospleniale nell’ippocampo dorsale e ventrale, nel subiculum e nella corteccia entorinale dopo il test di memoria recente (giorno 1) o remoto (30 giorni), con le corrispondenti microfotografie. L’espressione di c-fos è sovrapponibile a quella di Zif268 in tutte le regioni cerebrali tranne che nella corteccia retrospinale, dove solo i dati di Zif268 rivelano un coinvolgimento di quest’area.(Maviel et
Effetti dell’inattivazione neuronale della corteccia prefrontale, cingolata anteriore, posteriore e dell’ippocampo dorsale attraverso lidocaina (barre in nero) rispetto a un liquido cerebrospinale artificiale
(aCSF, barre in bianco), nel test di memoria recente (giorno 1) o remota (giorno 30). (Maviel et al., 2004). prefrontale, nella corteccia cingolata anteriore, nella corteccia cingolata posteriore, nella corteccia retrospleniale nell’ippocampo dorsale e ventrale, nel subiculum e nella orni), con le corrispondenti fos è sovrapponibile a quella di Zif268 in tutte le regioni cerebrali tranne che nella corteccia retrospinale, dove solo i dati di Zif268 rivelano un coinvolgimento di quest’area.(Maviel et
Effetti dell’inattivazione neuronale della corteccia prefrontale, cingolata anteriore, posteriore e cerebrospinale artificiale (aCSF, barre in bianco), nel test di memoria recente (giorno 1) o remota (giorno 30). (Maviel et al., 2004).
~ 29 ~
Infine per verificare se le modifiche funzionali fossero state accompagnate da cambiamenti strutturali, tra i quali sinaptogenesi e riorganizzazione laminare, è stato rilevato in aCC e PFC l’espressione della proteina GAP-43, proteina presinaptica, che controlla la crescita assonale e che viene utilizzata come marcatore della formazione di nuove sinapsi. Anche se non sistematicamente a 30 giorni è stato ritrovato un aumento della proteina, che potrebbe essere ritenuto responsabile della facilitazione e del potenziamento dell’LTP in quell’area. Anche Teixeira e collaboratori (2006) hanno cercato di sottolineare l’importanza della corteccia anteriore cingolata (aCC) in un compito spaziale, come quello del labirinto ad acqua di Morris, nell’espressione della memoria remota in ratti. Per valutare l’attivazione di tale area dopo il test di richiamo, hanno esaminato l’espressione del gene c-fos. In linea con le loro ipotesi gli autori riscontrarono un’espressione c-fos aumentata in aCC durante il test di memoria remota (30 giorni). Mentre nelle strutture ippocampali, CA1, CA3 e DG, l’espressione genetica è stata simile sia nel caso del richiamo a distanza sia per il richiamo recente (1 giorno). Anche se, l’attivazione dell’ippocampo si è rivelata indipendente dall’apprendimento, è stata significativamente maggiore rispetto ai controlli che non hanno ricevuto il training, indicando l’impiego di questa struttura nel task. Per confermare maggiormente queste dissociazioni funzionali tra le regioni cerebrali, anche in questo caso è stata effettuata l’inattivazione farmacologica con lidocaina prima del richiamo sia nella aCC che nell’ippocampo. Le performance dei soggetti del primo gruppo erano deficitarie nel test di richiamo tardivo e non nel recente, viceversa, il danno ippocampale provocato nel secondo gruppo ha effetti negativi su entrambe le prestazioni (Fig. 1-12).
~ 30 ~
Figura 0-12 Effetti dell’inattivazione con lidocaina di aCC e dHPC nella memoria spaziale recente (1 giorno) e
remota (30 giorni) testata dopo l’addestramento alla versione classica del water maze con la piattaforma nascosta. 10 minuti prima del test i soggetti ricevevano l’infusione di PBS, soluzione salina innocua (a), o lidocaina in aCC (b) o in dHPC (c). A sinistra, la percentuale del tempo speso nel cercare la zona target (T) comparata con le altre zone (O) durante il probe test. Al centro, plot della densità delle ricerche per ogni gruppo nei test recenti e remoti. Il colore della scala rappresenta il numero di passaggi per animale per 5x5 cm di area. A destra, tracciato di ricerca nel probe test remoto di un soggetto rappresentativo. La posizione della piattaforma è segnalata da un piccolo cerchio in basso a destra; inoltre un cerchio interno concentrico alla piattaforma indica l’area della vasca usata per quantificare il comportamento tigmotattico (Teixeira et al., 2006).
Questo studio è selettivamente incentrato sul ruolo di aCC nella memoria spaziale, ma, non bisogna dimenticare che altre aree giocano un ruolo fondamentale nel complesso network responsabile del consolidamento. Una di queste regioni è la corteccia entorinale, la quale funge da interfaccia di dialogo ippocampo-neocortex, la corteccia parietale e retrospinale, fondamentali rispettivamente per la rappresentazione di informazioni spaziali egocentriche e allocentriche (Mocovitch et al. 2005), e la corteccia infralimbica e prelimbica correlate con gli aspetti motivazionali della performance. A differenza di lesioni ad aCC e all’ippocampo, danni alle aree sopra citate portano solo a deficit spaziali minori.
~ 31 ~
Una strategia alternativa potrebbe essere di interrompere il trasferimento della traccia, mediante una lesione che va a bloccare il dialogo tra l'ippocampo e la corteccia poco dopo il training, in modo da permettere la normale formazione di tracce mnemoniche ippocampali ma impedire il loro ulteriore consolidamento nelle reti corticali. Ad esempio, un danno alla proiezione che dallo strato III della corteccia entorinale arriva alla regione CA1 dell'ippocampo, consente una normale funzionalità ippocampale ma impedisce il dialogo ippocampo-corteccia. Ratti con simili lesioni, sottoposti al labirinto di Morris, mostrano un apprendimento spaziale normale e delle prestazioni ottime in seguito al test di memoria recente ,ma la memoria è compromessa se testata in seguito a un intervallo di ritenzione maggiore. Ciò indica che le interazioni cortico-ippocampali sono necessarie per la formazione di memoria spaziale remota (Remondes & Shuman, 2004). La corteccia cingolata anteriore aCC, insieme alla corteccia prelimbica e infralimbica, fanno parte della corteccia mediale prefrontale (mPFC), regione fortemente interconnessa al suo interno e al suo esterno con aree sensoriali (Teixeira et al., 2006). Recenti studi mirati all’individuazione dei net-work che sottendono i processi mnestici, hanno dimostrato che alcuni nuclei talamici anteriori compresi alcuni nuclei intralaminari , svolgono un ruolo selettivo nei processi di memoria, grazie alle connessioni dirette e indirette con il sistema ippocampale e con la corteccia prefrontale, partecipano alla riorganizzazione tempo-dipendente delle tracce di memoria durante il consolidamento. Infatti, lesioni a queste aree influenzano l'acquisizione ed il recupero di tracce di memoria spaziale acquisite in seguito al training nel ratto. Questo studio fornisce la prima evidenza del ruolo
della regione talamica come componente fondamentale della rete
extraippocampale coinvolte nella riorganizzazione tempo-dipendente della memoria spaziale nel corso del consolidamento (Lopez et al., 2009) ed in particolare mette in luce alcuni percorsi presenti tra l’ippocampo e la mPFC essenziali nella riorganizzazione tempo-dipendente delle tracce di memoria. Uno dei percorsi che collega l’ippocampo direttamente alla corteccia mediale prefrontale, è originario dalla regione CA1 dell'ippocampo e dal subiculum, e termina nella corteccia prelimbica, mentre i nuclei talamici sono coinvolti in due