Capitolo 1
Anatomia
e
fisiologia
della
respirazione
L’apparato respiratorio è un insieme di organi e tessuti che servono per esplicare la funzione respiratoria; è costituito dalle vie aere superiori ed inferiori e permette lo scambio di gas tra aria ambiente e sangue (Roberts, 2000).
1.1 Strutture anatomiche: definizioni
L’apparato respiratorio si estende dalla bocca e dal naso fino agli alveoli (Roberts, 2000).
L’aria viene inspirata attraverso le narici, dove è sottoposta ad una prima filtrazione dalle particelle estranee presenti, ed arriva alla faringe passando attraverso i meati nasali, dove viene riscaldata ed arricchita di vapor acqueo, quindi passa dalla laringe nella trachea, arriva ai bronchioli, ai dotti alveolari ed agli alveoli, circondati dai capillari polmonari. Uno strato epiteliale monocellulare divide l’aria contenuta negli alveoli dal sangue
contenuto nei capillari. La superficie alveolare in contatto con i capillari è estremamente estesa (Roberts, 2000).
Ciascun polmone è circondato da una pleura viscerale ed una pleura parietale. La pleura viscerale è una membrana liscia, umida e sottilissima che riveste ciascun polmone; la pleura parietale, di costituzione simile alla pleura viscerale, da un lato è attaccata alla cavità toracica e dall’altro lato è adesa alla pleura viscerale. L’adesione tra le due pleure è resa possibile dalla presenza di un sottilissimo film liquido tensio-attivo interposto tra le due membrane, il surfactante.
Quando la cavità toracica si espande, la pleura parietale segue le pareti interne del torace a cui è attaccata e trascina con sé la pleura viscerale; in questo modo anche il polmone si espande. (De Rensis, 2001).
1.2 Ciclo respiratorio
L’attività respiratoria è caratterizzata da una fase inspiratoria, durante la quale dell’aria viene introdotta nelle vie respiratorie, ed una fase espiratoria che serve invece per espellerla.
La fase inspiratoria è un processo attivo determinato dalla contrazione dei muscoli inspiratori. Il principale di tutti è il diaframma. Quando questo è rilassato protende nella cavità toracica riducendone il volume, quando è contratto si appiattisce, espandendo la cavità toracica. Gli altri muscoli coinvolti sono i muscoli intercostali, interni ed esterni, che contraendosi sollevano le coste ed aumentano il diametro toracico ( De Rensis, 2001).
Ci sono poi dei muscoli accessori della respirazione (sternocleidomastoidei e scaleni) che diventano importanti durante l’esercizio fisico o durante le affezioni respiratorie (Roberts, 2000).
Durante l’inspirazione, quando il torace si espande, la pressione intrapleurica varia da –2,5 mmHg (in rapporto alla pressione atmosferica) a –6 mmHg (De Rensis, 2001). Nelle fasi iniziali dell’inspirazione il volume dei polmoni aumenta, ma non la quantità di aria al loro interno: questo comporta un aumento della pressione negativa intrapolmonare che causa un flusso di aria verso l’interno dei polmoni.
Quando la pressione intrapolmonare è di nuovo uguale a quella atmosferica questo flusso di aria termina e così anche la fase inspiratoria.
La fase espiratoria è invece un processo passivo, che non richiede cioè l’intervento di una attività muscolare, ma è determinato solamente dalla retrazione elastica dei polmoni.
Durante l’espirazione il volume della cavità toracica si riduce e inizialmente l’aria contenuta nei polmoni viene compressa per cui la pressione intrapolmonare aumenta e supera quella atmosferica: a questo punto l’aria viene espulsa dai polmoni fino a quando non eguaglia la pressione atmosferica. La fase espiratoria termina quindi quando la pressione dell’aria nei bronchi è uguale a quella atmosferica.
Generalmente i cicli respiratori sono continui (Reece, 2002). Cicli respiratori complementari sono caratterizzati da profonde e rapide inspirazioni seguite da espirazioni di durata più lunga. Questo tipo di cicli viene frequentemente
chiamato sospiro. Quando si verifica è probabilmente un meccanismo di compensazione in caso di un’insufficiente ventilazione.
Il respiro addominale predomina durante la respirazione normale e tranquilla. Questo tipo di respiro è caratterizzato da visibili movimenti addominali causati dalla compressione del diaframma. Durante l’espirazione l’addome retrocede.
Il respiro costale è caratterizzato da marcati movimenti costali; quando la respirazione comincia da essere difficoltosa o in condizioni di algia dell’addome questo tipo di respiro diventa predominante. (Reece, 2002).
Prima di raggiungere gli alveoli l’aria deve passare attraverso il naso o la bocca, la faringe, la trachea, i bronchi ed i bronchioli. Poiché l’ aria presente in queste strutture non può partecipare allo scambio gassoso, che avviene solo a livello alveolare, questo spazio è detto spazio morto anatomico. Nei soggetti normali coincide con lo spazio morto fisiologico, rappresentato dal volume di gas che è inspirato ma che non partecipa agli scambi respiratori nelle vie aeree e negli alveoli (West, 2005).
Nei soggetti con alveoli anche parzialmente non funzionanti oppure con abnorme rapporto tra flusso ematico e funzione polmonare ci può essere invece una differenza anche di dieci volte superiore tra quello fisiologico e quello anatomico.(Guyton, 1983).
Il volume di aria che passa attraverso la bocca od il naso durante ciascun atto inspiratorio ed espiratorio è detto volume tidalico. Il volume occupato dallo spazio morto è circa 1/3-1/4 del volume tidalico.
Il volume di aria che raggiunge gli alveoli (ventilazione polmonare), disponibile per gli scambi gassosi, dipende dal volume tidalico, lo spazio morto e la frequenza respiratoria secondo il seguente rapporto:
Ventilazione alveolare = (vol. tidalico–spazio morto) x frequenza respiratoria
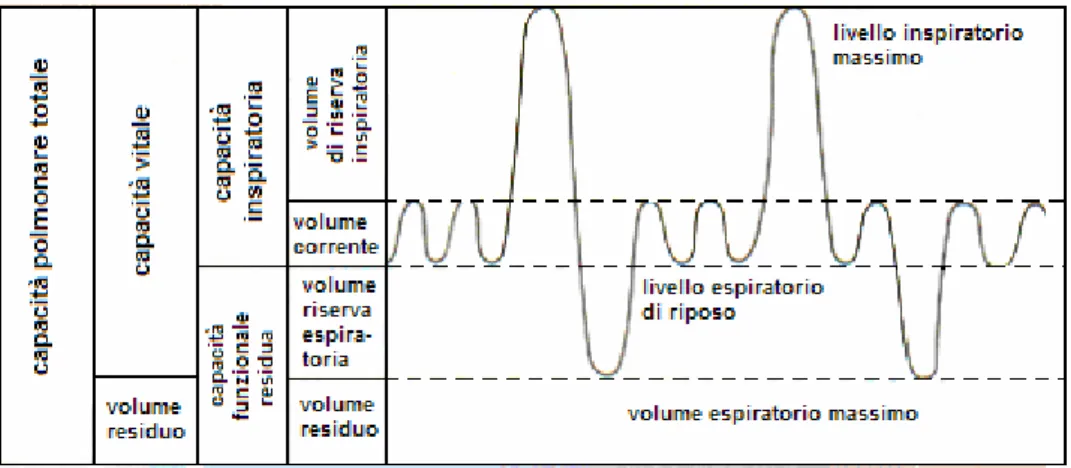
Altri volumi respiratori sono (Fig.1.1):
Il volume inspiratorio di riserva: la quantità di aria in più che può essere introdotta mediante una inspirazione forzata;
Il volume espiratorio di riserva: la quantità di aria che può essere espulsa mediante una espirazione forzata;
Il volume residuale: la quantità di aria che rimane anche dopo un’espirazione forzata e che permette al polmone di non collassare.(De Rensis, 2001)
La capacità inspiratoria è la somma del volume corrente (o tidalico) e del volume di riserva inspiratorio: è la quantità di aria che un soggetto può inspirare dalla fine di un’espirazione tranquilla sino alla distensione massima dei suoi polmoni.
La capacità funzionale residua è uguale alla somma del volume corrente, volume di aria inspirata o espirata ad ogni atto respiratorio tranquillo, e del
volume residuale. È rappresentato quindi dalla quantità di aria che residua nel polmone alla fine di una espirazione normale.
La capacità vitale è la somma del volume di riserva inspiratoria, del volume corrente e del volume di riserva espiratoria. È la massima quantità di aria che un soggetto può espellere dai suoi polmoni con un’espirazione forzata al massimo al termine di un’inspirazione forzata anch’essa al massimo. La capacità polmonare totale è il massimo volume di aria che può essere contenuto nei polmoni al termine di un’inspirazione forzata.(Guyton, 1983)
Figura 1.1: Volumi e capacità polmonari
1.3
Regolazione del respiro
Il sistema nervoso centrale è in grado di regolare il ritmo ventilatorio in modo da far fronte alle esigenze dell’organismo e cercare di mantenere i livelli di pressione parziale dell’ossigeno (PO2) e di anidride carbonica (PCO2) nel sangue costanti anche in corso di attività muscolare intensa (De Rensis, 2001).
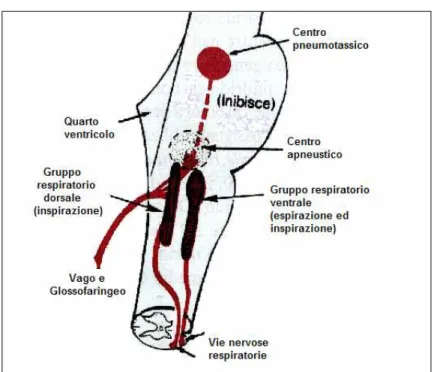
Il centro respiratorio è costituito da un gruppo diffusamente sparso di neuroni, situati bilateralmente nella formazione reticolare del bulbo e del ponte (Fig. 1.2). Consta di tre formazioni principali:
• Il centro bulbare della ritmicità • Il centro apneustico
• Il centro pneumotassico
Il centro bulbare della ritmicità sembra essere il più importante dei tre, è detto anche centro respiratorio bulbare e si trova nella formazione reticolare del bulbo.
I neuroni inspiratori ed i neuroni espiratori sono entrambi localizzati nel centro bulbare. Quando la respirazione si intensifica sia i neuroni inspiratori che quelli espiratori diventano molto più attivi e trasmettono un numero più grande di segnali. È il centro respiratorio bulbare che stabilisce il ritmo basale della respirazione.
Nelle normali condizioni di riposo l’inspirazione dura la metà dell’espirazione con cicli respiratori variabili tra 20 e 35 al minuto (Aguggini, 1998). Questi diminuiscono od aumentano in proporzione in base all’attività del soggetto. Tuttavia il centro da solo non è in grado di assicurare un tipo di respiro normale e scorrevole.
Al centro respiratorio bulbare pervengono segnali afferenti, oltre che dal midollo spinale, anche dalla corteccia cerebrale e dal mesencefalo, dal centro pneumotassico, nella parte superiore del ponte, e da quello apneustico, nella parte inferiore del ponte. Tutti questi segnali influiscono sul ritmo respiratorio e contribuiscono a realizzare un tipo di respirazione normale e scorrevole.
Il centro apneustico e quello pneumotassico si trovano nella formazione reticolare del ponte. Essi non sono indispensabili per il mantenimento del ritmo respiratorio basale (Guyton, 1983).
Si pensa che il centro pneumotassico regoli la sensibilità del centro respiratorio agli stimoli che sollecitano la fine dell’inspirazione e facilitano l’espirazione.
Il centro apneustico sembra essere invece coinvolto nelle inspirazioni profonde (apneusi), e pare che i respiri complementari (sospiri) siano manifestazioni della sua attività (Reece, 2002).
Oltre al controllo neuronale sul centro del respiro ci sono anche sostanze chimiche del sangue che modificano il ritmo respiratorio basale.
Visto che queste sostanze sono presenti nel sangue, questo meccanismo viene definito controllo umorale.
In maniera specifica gli agenti chimici cui viene fatto riferimento sono anidride carbonica, ossigeno e ioni idrogeno. La loro concentrazione nel sangue arterioso modifica la ventilazione alveolare in questo modo:
• Un aumento della pressione parziale dell’anidride carbonica causa aumento della ventilazione alveolare; una sua diminuzione causa, viceversa, diminuzione della ventilazione alveolare.
• Un aumento della concentrazione di ioni idrogeno causa un incremento della ventilazione alveolare; un abbassamento di concentrazione fa diminuire la ventilazione alveolare.
• Un abbassamento della pressione parziale di ossigeno determina aumento della ventilazione alveolare; un suo aumento causa invece una diminuzione della ventilazione alveolare (Reece, 2002).
Le aree chemiosensibili della superficie ventrale del bulbo hanno un’elevata sensibilità per le variazioni di concentrazione di ioni idrogeno del liquido
interstiziale del cervello. I chemiocettori di queste aree eccitano il centro respiratorio, causando aumento del volume tidalico e della frequenza.
Mentre gli ioni idrogeno diffondono con difficoltà attraverso la barriera sangue-liquido cerebrospinale e la barriera emato-encefalica, l’anidride carbonica diffonde liberamente ed esercita indirettamente la sua influenza sulla ventilazione attraverso gli intermedi della sua reazione di idratazione cioè gli ioni idrogeno (CO2+H2O H2CO3 H++HCO3-). Quindi se la concentrazione di anidride carbonica nel sangue dovesse aumentare, la PCO2 del liquido interstiziale del bulbo e del liquido cerebrospinale aumenterebbe, inducendo la formazione di H+ dalla reazione di idratazione (Reece, 2002).
La maggior parte della stimolazione del centro del respiro proviene dalle variazioni di H+ del liquido interstiziale. Tuttavia poiché i chemiocettori sono localizzati vicino alla superficie del bulbo, anche la diffusione di H+ dal liquido cerebrospinale ai chemiocettori è considerata fonte di stimolazione. È importante sottolineare che l’accumulo di anidride carbonica e ioni idrogeno, prodotti dalle cellule, incrementa la ventilazione per favorirne l’eliminazione e una riduzione della quantità delle sostanze consumate (ossigeno) incrementa parimenti la ventilazione per supplire alla loro deplezione.
Anche la diminuzione di ioni idrogeno e di anidride carbonica riducono la ventilazione; così facendo se ne previene l’eccessiva perdita che altrimenti
causerebbe drastiche modificazioni del pH del liquido corporeo (Reece, 2002).
In pratica l’iperventilazione può abbassare la concentrazione di anidride carbonica e di conseguenza di H+ fino al punto di provocare un’alcalinizzazione dei liquidi dell’organismo. Questo effetto, tramite il quale le ridotte concentrazioni di anidride carbonica e di ioni idrogeno riducono la ventilazione alveolare, è conosciuto come effetto braking (effetto freno). Esso può essere osservato entro certi limiti anche per l’aumento di concentrazioni di ossigeno, ma tale effetto è assai ridotto.
Le interazioni tra anidride carbonica, ossigeno e ioni idrogeno sono molto evidenti in alcune condizioni. È possibile, per mancanza di ossigeno, ottenere un incremento della ventilazione fino al punto in cui una diminuzione di anidride carbonica e ioni idrogeno in seguito ad iperventilazione dà luogo all’effetto braking; questo previene la grave alcalosi respiratoria che altrimenti potrebbe svilupparsi. Tale situazione si determina nelle ascensioni ad altitudine elevata, dove la PO2 è ridotta. L’effetto braking dell’anidride carbonica e degli ioni idrogeno si manifestano i primi giorni, poi l’adattamento ne riduce l’influenza cosicché la ventilazione aumenta per compensare la diminuzione dell’ossigeno atmosferico. Parte dell’adattamento è collegata alla minore risposta dell’effetto braking dell’anidride carbonica e degli ioni idrogeno (Reece, 2002).
L’ossigeno non è in grado di influenzare direttamente le aree chemiosensibili del bulbo; una diminuzione della concentrazione
dell’ossigeno viene avvertita da recettori periferici che si trovano nei corpi carotidei ed aortici. Queste entità anatomiche conosciute come glomi carotideo ed aortico si trovano per la precisione alla biforcazione delle carotidi e nell’arco aortico.
Essi rilevano i cambiamenti delle pressioni parziali di anidride carbonica ed ossigeno, le variazioni di ioni idrogeno ed influiscono sul centro respiratorio attraverso la trasmissione di impulsi che viaggiano con le fibre afferenti dei nervi glossofaringeo (glomi carotidei) e vago (glomi aortici). Sebbene il bulbo sia la sede principale per il rilevamento dei cambiamenti di concentrazione di CO2 e H+, è stato dimostrato che questi chemiocettori periferici forniscono il 30% degli stimoli ventilatori in risposta alle variazioni di PCO2.
La disposizione anatomica dei chemiocettori del glomo carotideo ed aortico è tale da permettere il rilievo della pressione parziale dell’ossigeno. Questi piccoli organi sono abbondantemente perfusi di sangue e l’informazione sull’ossigeno necessario all’attività basale è ottenuta dall’ossigeno disciolto (Reece, 2002).
Il centro respiratorio del cane non risponde soltanto agli usuali stimoli, ma anche alla temperatura corporea interna.
L’integrazione di questi impulsi permette al centro respiratorio di rispondere alle necessità metaboliche regolando la ventilazione dello spazio morto respiratorio. Quest’ultima nell’ansito aumenta, il che contribuisce al
raffreddamento corporeo tramite l’evaporazione d’acqua dalle mucose dei tessuti coinvolti (Reece, 2002).
Per far espandere i polmoni i muscoli devono spendere energia, ciò costituisce una parte del “lavoro” respiratorio. Oltre al lavoro richiesto per espandere semplicemente i polmoni ne serve altro per vincere due fattori che contrastano l’espansione e la costrizione polmonare: la viscosità del tessuto polmonare, detta anche resistenza tessutale non elastica e la resistenza delle vie aeree (Guyton, 1983).
Per la normale respirazione l’energia richiesta è molto piccola, ma quando le vie aeree si restringono, come ad esempio nell’asma e nell’enfisema ostruttivo, la resistenza delle vie respiratorie può essere tanto grande da richiedere ai muscoli respiratori una quantità di energia in più semplicemente per forzare l’aria lungo le vie aeree stenotiche (Guyton, 1983).
In termini di percentuale il costo energetico per la ventilazione polmonare durante una respirazione tranquilla ammonta a circa il 2-3% dell’energia spesa complessivamente dall’intero organismo.
Le malattie dei polmoni, che ne fanno aumentare la viscosità o la resistenza, fanno aumentare il lavoro respiratorio fino ad impegnare 1/3 della spesa energetica totale (Guyton, 1983).
1.4 Ematosi
La quantità di aria alveolare che viene sostituita con aria atmosferica nuova è, per ogni atto respiratorio, solo un settimo del totale, ci vogliono quindi numerosi atti respiratori per rinnovare la maggior parte dell’aria alveolare; questo lento rinnovamento è importante perché evita brusche variazioni delle concentrazioni dei gas nel sangue prevenendo eccessivi aumenti o diminuzioni dell’ossigenazione dei tessuti o una eccessiva concentrazione di anidride carbonica o pH tessutale in caso di temporanea interruzione del respiro (Guyton, 1983).
L’ossigeno viene diffuso continuamente nel sangue polmonare mentre nuovo ossigeno perviene negli alveoli, quanto più rapidamente l’ossigeno diffonde nel sangue, tanto più bassa si fa la sua concentrazione negli alveoli.
La concentrazione di ossigeno negli alveoli è controllata in primo luogo dalla velocità con la quale l’ossigeno viene assorbito dal sangue e in secondo luogo dalla velocità con cui il nuovo ossigeno entra nei polmoni dall’esterno. Nell’organismo viene prodotta continuamente anidride carbonica, che viene poi riversata negli alveoli e da questi rimossa tramite la ventilazione.
I due fattori che condizionano il valore della pressione parziale di anidride carbonica (PCO2) nei polmoni sono la velocità di diffusione dell’anidride carbonica dal sangue negli alveoli e la velocità con la quale la ventilazione alveolare riesce a rimuovere l’anidride carbonica dagli alveoli.
L’entità della PCO2 aumenta in modo direttamente proporzionale con la velocità di escrezione dell’anidride carbonica e si abbassa in modo inversamente proporzionale alla ventilazione alveolare. Perciò la concentrazione dell’ossigeno e quella dell’anidride carbonica sono condizionate dalla velocità di assorbimento e, rispettivamente, di escrezione di questi due gas, nonché dalla ventilazione alveolare (Guyton, 1983).
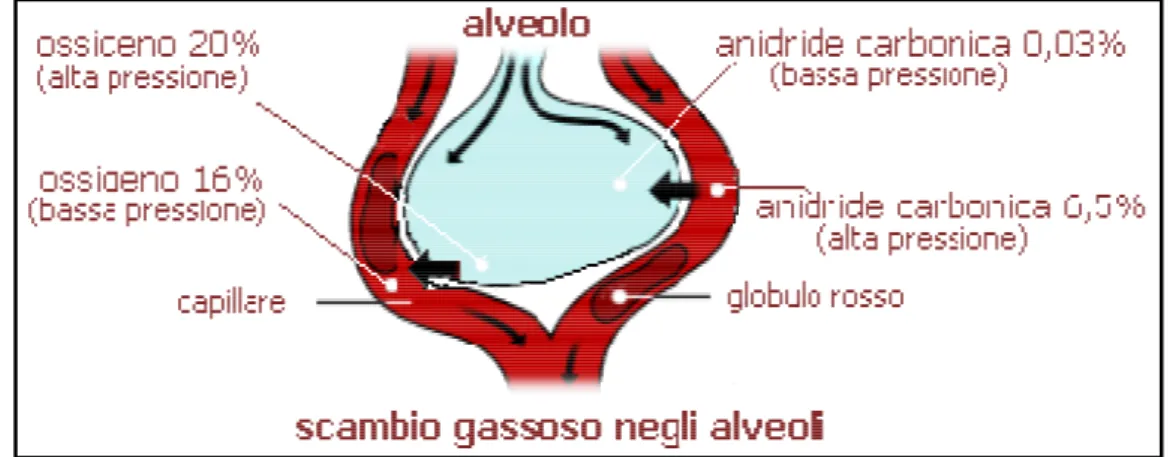
Tutti gli spostamenti del gas alveolare nel polmone avvengono per diffusione passiva; un gas si muove da una regione all’altra solo quando la pressione parziale del gas è più grande in una regione rispetto all’altra (Fig. 1.3) (Aguggini, 1998).
Figura 1.3: Scambio alveolare di ossigeno ed anidride carbonica
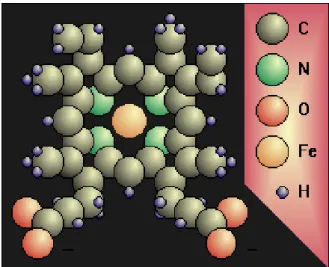
L’ossigeno è trasportato dalle molecole di emoglobina, una cromoproteina contenuta nei globuli rossi (Fig. 1.4).
Essa è formata da una parte proteica e da un pigmento, l’eme. A sua volta la parte proteica è composta da 4 catene polipeptidiche ognuna delle quali contiene un gruppo eme. Ogni gruppo eme contiene un atomo di ferro allo
stato ferroso che si combina debolmente e reversibilmente con l’ossigeno. Quindi una molecola di emoglobina può trasportare 4 molecole di ossigeno.
Fig 1.4.: Molecola di emoglobina; C, carbonio; N, azoto; O, ossigeno; Fe, ferro; H, idrogeno
L’ossigeno che viene consumato dalle cellule del corpo proviene dall’ossigeno disciolto nell’ambiente interno (liquido interstiziale). A sua volta l’ossigeno interstiziale viene rimpiazzato con l’ossigeno disciolto nel plasma e questo se ne rifornisce prendendolo dagli eritrociti.
Quando il sangue povero di ossigeno arriva ai polmoni, la situazione si capovolge, l’ossigeno diffonde dagli alveoli al liquido interstiziale e da questo passa al plasma ed agli eritrociti del circolo sanguigno e, negli eritrociti si lega all’emoglobina (Reece, 2002).
L’ossigeno e l’anidride carbonica hanno una bassa solubilità nei liquidi, la maggioranza dell’ossigeno è trasportato legato all’emoglobina nei globuli rossi mentre l’anidride carbonica in forma di ioni bicarbonato o come carbamino-emoglobina.
La quantità di ossigeno che l’emoglobina riesce a trasportare viene diminuita dalla presenza dell’anidride carbonica, dall’aumento dell’acidità e della temperatura del sangue e dalle concentrazioni di fosfati organici presenti nei globuli rossi ( De Rensis, 2001).
L’anidride carbonica prodotta dai tessuti promuove il distacco dell’ossigeno dall’emoglobina, distacco dovuto alla maggior affinità di questa per la molecola di emoglobina stessa e dalla formazione di carbamino-emoglobina oltre che dall’aumento di acidità del sangue.
L’acidità facilita il distacco dell’ossigeno dall’emoglobina in modo da favorire i tessuti durante l’esercizio fisico quando aumenta la produzione di acido lattico e si abbassa il pH ( De Rensis, 2001).
Anche l’aumento della temperatura favorisce il distacco dell’ossigeno dall’emoglobina in modo che i tessuti che hanno un metabolismo più elevato, e quindi una temperatura più elevata, e un maggior fabbisogno di ossigeno ne possano ricevere una quantità maggiore.
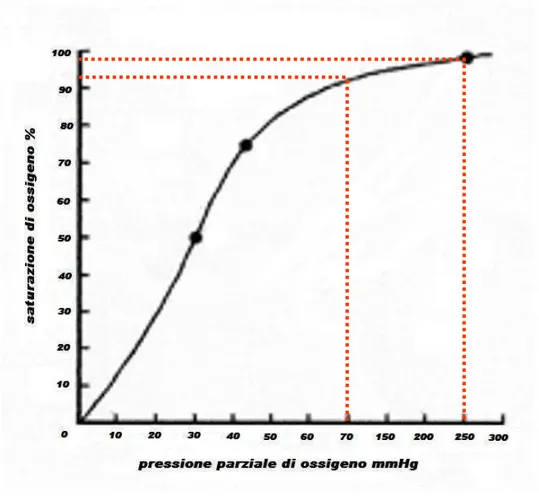
La curva di dissociazione dell’emoglobina (Fig. 1.5) ha una forma iperbolica dove la parte più ripida tra i 20-60 mmHg di PO2 indica che l’ossigeno è rapidamente ceduto ai tessuti.
Figura 1.5: Curva di dissociazione dell’emogobina
La parte più piatta sopra i valori di 80 mmHg indica che il sangue arterioso ha un contenuto costante di ossigeno anche quando la composizione dell’aria alveolare varia molto: queste caratteristiche facilitano la saturazione dell’emoglobina quando il sangue è a contatto con gli alveoli e allo stesso tempo fa si che l’ossigeno venga ceduto ai tessuti con una bassa PO2 (De Rensis, 2001).
Durante una respirazione tranquilla, il sangue è gia pressoché saturo di ossigeno quando ha percorso il primo terzo della lunghezza del capillare, e solo una piccola parte di quantità di ossigeno in più vi entra negli ultimi due
terzi del percorso. Pertanto, anche se durante il lavoro muscolare risulta abbreviato il tempo di esposizione del sangue, questo può tuttavia ossigenarsi in modo completo (Guyton, 1983).