RIASSUNTO
Parole chiave: Piccione, urine, plasma, SDS-PAGE, istologia renale
Il piccione (Columba livia) rappresenta una delle specie sinantrope più diffuse, ma allo stesso tempo è abitualmente allevato per la carne, come uccello ornamentale o come piccione viaggiatore. Questo uccello, proprio per la sua grande distribuzione, è frequentemente ospitato nei centri di recupero. E’ un animale rustico, ma facile da gestire, di piccola–media taglia, per questo rappresenta un modello per effettuare studi sugli uccelli.
In questo lavoro sono stati presi in esame 34 esemplari di piccione, ospitati in un centro di recupero. Da ciascun animale sono state raccolte le urine, il plasma e un frammento di parenchima renale. Per due di questi soggetti è stato raccolto anche un campione di urina ureterica. Il plasma e le urine, sia quelle cloacali che ureteriche, sono stati analizzati con la metodica SDS-PAGE.
Per 26 soggetti, è stato effettuato anche un esame chimico delle urine con spettrofotometro, per il dosaggio delle proteine urinarie.
Il tessuto renale è stato preparato per l’istologia e i referti confrontati con i risultati elettoforetici. Le bande ottenute con l’elettroforesi delle urine ureteriche sono risultate le stesse nei due soggetti sebbene il rene di uno di loro fosse sano e l’altro risultasse patologico all’istologia. I risultati elettroforetici delle urine cloacali, messi a confronto con quelle ureteriche, evidenziano a seconda dei casi la presenza di ulteriori bande o l’assenza di alcune di esse.
SUMMARY
Key-words: Pigeon, urine, plasma, SDS-PAGE, renal histology
Pigeon (Columba livia) is one of the most widespread synanthropic avian species and it is usually used for its meat or as ornamental bird and carrier-pigeon. This avian species, because of his widespread distribution, it is frequently rescued by wildlife rescue centers. Pigeon is a rustic animal, easy to manage and small-medium sized, so it represent a model for avian’s studies.
In this study 34 pigeons hospitalized in a wildlife rescue center have been examined.
Urine, plasma and renal tissue was collected from each animal. Ureteric urine was also collected from 2 animals. Plasma and ureteric and cloacal urine were analyzed with SDS-PAGE.
In 26 pigeon urine chemical analysis with spectrophotometer for urinary proteins assay was performed. Renal tissue was submitted to histology and reports compared with electrophoresis results.
The bands from ureteric urine electrophoresis were the same in both subjects although one kidney was healthy and the other one was patholologic at the histology.
INTRODUZIONE
Il piccione originariamente era presente soprattutto nell’Europa meridionale e nei paesi che si affacciano sul mar mediterraneo, ma attualmente ad opera dell’uomo risulta essere ubiquitario.
Possiede una buona capacità di adattamento che gli ha permesso con il passare del tempo di colonizzare vaste aree, compreso l’ambiente urbano.
I piccioni presenti in città non possiedono più le caratteristiche dei soggetti ancestrali, poiché si sono ampiamente incrociati con soggetti di allevamento inselvatichiti.
L’elevato numero di piccioni presenti nelle aree urbane e non, fa si che questi siano spesso coinvolti in incidenti o trovati in cattivo stato di salute da persone che li conducono in centri di recupero. In questi centri gli animali vengono curati al fine di liberarli nuovamente nell’ambiente di origine, una volta completamente guariti.
Il rene degli uccelli, al pari di quello dei mammiferi rappresenta un organo emuntore, coinvolto nell’omeostasi dei liquidi e nell’eliminazione dei cataboliti.
Esistono numerose differenze fra l’apparato urinario degli uccelli e dei mammiferi, ma due sono quelle fondamentali; gli uccelli sono uricotelici, cioè eliminano i prodotti azotati sotto forma di acido urico e non possiedono una vescica urinaria, quindi le secrezioni provenienti da apparato digerente, urinario e genitale si mischiano nella cloaca.
Gli uccelli, sia i domestici che i selvatici, al pari dei mammiferi, sono frequentemente interessati da patologie urinarie, che probabilmente vengono spesso sottovalutate.
La diagnosi di queste patologie risulta spesso difficile, l’unico modo per fare una diagnosi certa è attraverso la biopsia renale con successiva istologia.
La raccolta di urine pure necessita di particolari accortezze negli uccelli, dato che possiedono la cloaca.
L’analisi chimico-fisica delle urine dà spesso risultati dubbi, inoltre non tutte le tecniche utilizzate nei mammiferi possono essere applicate alle urine degli uccelli che contengono acido urico.
Con questo lavoro ci siamo proposti l’obiettivo di approfondire le tecniche di collezione di campioni urinari ed ematici, anestesiologiche, endoscopiche e bioptiche su 34 piccioni presenti al CRUMA (Centro di Recupero Uccelli Marini ed Acquatici) di Livorno al fine
di verificare l’applicabilità delle metodologie diagnostiche per la patologia urinaria conosciute nei mammiferi anche in questa specie animale.
1. IL PICCIONE (Columba livia)
1.1 TASSONOMIA
Dominio: Eukaryota Regno: Animalia Sottoregno: Eumetazoa Superphylum: Deuterostomia Subphylum: Vertebrata Superclasse: Tetrapoda Classe: Aves Sottoclasse: Carinatae Infraclasse: Neornithes Ordine: Columbiformes Famiglia: Columbidae Sottofamiglia: Columbinae Genere: Columba Specie: liviaNome sientifico: Columba livia
Dal Colombo selvatico (Columba livia livia) sono derivati i piccioni che popolano le nostre città e le numerose varietà di colombi domestici. Altri colombi selvatici presenti in Italia e in Europa sono il Colombaccio (Columba palumbus) e la Colombella (Columba oenas).
1.2 DISTRIBUZIONE
Il piccione è tipico dell'Europa meridionale, del nord Africa, e del Medio Oriente.
La sua distribuzione originaria comprendeva gran parte dell'Europa meridionale, Scozia, Irlanda, Africa del Nord, India, Palestina e Siria; in seguito si è diffuso soprattutto per opera dell'uomo tanto da diventare ubiquitaria. Oggi la specie ancestrale si è ibridata con le molte razze domestiche e le poche popolazioni pure vivono nelle isole Ebridi e Shetland, al largo della Scozia e lungo alcune coste della Sardegna e della Spagna.
Il Piccione, è una delle specie di columbidi più diffusa in Italia soprattutto nelle grandi città, colonizza edifici e sottotetti .
Tra le specie sinantrope il colombo (Columba livia) è una di quelle che ha conosciuto in questi ultimi anni un vero e proprio boom demografico, raggiungendo in alcune aree del nostro paese, la densità media di 3000 individui/km2, con un numero di colombi che supera le 100.000 unità nelle grandi città (Sbragia et al., 2001).
1.3 ASPETTI MORFOLOGICI
Il colombo (Columba livia), è lungo 30-35 cm con apertura alare di 62-68 cm, il peso oscilla tra 180 - 370 grammi. La testa, il collo e la parte superiore del petto sono grigio blu scuro nell'adulto con riflessi iridescenti viola e verde smeraldo. Gli occhi sono arancioni o rossastri, circondati da anelli grigio-bianco negli adulti, mentre nei giovani con meno di 6 - 8 mesi gli occhi sono di colore marrone. Il becco è scuro, il colore delle zampe invece va dal rosso al grigio. Il dorso, le ali e il ventre sono grigie chiare. Le ali presentano due bande scure formate dalle estremità delle penne secondarie e di quelle che proteggono e coprono le secondarie, mentre le primarie sono di colore grigio scuro. La groppa è bianca, la coda grigia chiara con una fascia terminale nera.
Sebbene il piumaggio originario del piccione fosse grigio, attualmente in seguito all’incrocio con individui di allevamento che hanno riacquistato la libertà, sono presenti numerose varietà di piumaggio sia per quanto riguarda la colorazione: grigio,castano, nero o bianco; sia la livrea: barrata, uniforme, a mosaico.
È un animale molto resistente e veloce nel volo. La vita di un piccione comune varia dai 3 ai 5 anni allo stato selvatico, ma i soggetti addomesticati possono raggiungere anche 15-17
Non è presente dimorfismo sessuale evidente tra il maschio e la femmina, solo piccole differenze come profilo della testa più dolce, becco più fino ed una silhouette del corpo più snella nella femmina. I due sessi sono invece facilmente distinguibili quando stanno insieme, dall’atteggiamento e dai richiami fatti durante il corteggiamento.
1.4 BIOLOGIA
Si tratta di una specie non migratrice, fortemente gregaria, ciba prevalentemente di granaglie sia coltivate che spontanee, integrando la dieta con bacche e piccoli invertebrati come lumache o chiocciole.
Nelle aree urbane gran parte del cibo proviene dall'uomo (pane, pasta, briciole, rifiuti), spesso fornito intenzionalmente dai cittadini zoofili.
Alcuni gruppi di colombi si spostano nelle campagne circostanti, soprattutto durante le semine ed i raccolti, effettuando i cosiddetti "voli di foraggiamento", fino a circa 10 km dalle periferie (Baldaccini et al,1989).
Volano di solito in coppia o a piccoli gruppi fino a formare grandi stormi per la ricerca del cibo.
Raggiungono la maturità sessuale a 4-6 mesi. Il maschio corteggia la femmina tubando e gonfiando il collo mentre la femmina si fa rincorrere. Sono monogami, e la coppia rimane la stessa per tutta la vita.
Solitamente costruiscono il nido sulle sporgenze di costruzioni coperte (es. tetti), servendosi di piccoli ramoscelli. Il maschio porta alla compagna il materiale necessario alla costruzione, e la femmina costruisce il nido. La femmina depone due uova di colore bianco, che vengono covate per 17-18 giorni da entrambi i genitori che si alternano di giorno la femmina e di notte il maschio.
I piccioncini alla nascita sono ciechi, coperti di scarso piumino e per i primi 10 giorni di vita vengono alimentati con una sostanza secreta dal gozzo dei genitori, detta “latte di gozzo”; successivamente l'alimentazione sarà costituita oltre che dal secreto,anche dalle granaglie ingerite dai genitori. Il piumino inizia a cadere dopo 10-12 giorni gradualmente sostituito dalle penne definitive; il piumaggio definitivo è completo a circa 3 settimane. In un mese i piccoli sono pronti per volare ed abbandonare il nido. In allevamento la coppia può fare anche 6-7 nidiate in un anno (Balasini, 2001).
Il successo e le possibilità di espansione dipendono, oltre che dal buon adattamento all’ambiente urbano, anche dallo scarso numero di predatori che incontrano nelle città. I loro nemici naturali sono rapaci, come il falco pellegrino, astore, sparviero ed allocco, tutti uccelli raramente visibili in città e che quindi esercitano uno scarso controllo su questa specie. Uova e piccoli sono invece più esposti al pericolo di predazione da parte di topi, ratti, faine e cornacchie. Questi animali sono in grado di esercitare una certa pressione di selezione,contribuendo a limitare le possibilità riproduttive del colombo.
1.5 HABITAT
Il colombo selvatico frequenta le zone rocciose marine e gli ambienti carsici o comunque rocciosi dell’entroterra, dove sono presenti cavità e sporgenze riparate che utilizza sia per nidificare che per proteggersi.
Il colombo torraiolo è molto comune nelle aree urbane e negli ambienti agricoli.
Presente in quasi tutte le città italiane ed in molte europee, popola soprattutto le piazze ed i parchi. E' in grado di tollerare le situazioni più antropizzate, quali industrie, stazioni ferroviarie, sottopassaggi di metropolitane, mercati, ecc.
Per una qualsiasi specie di uccello, la capacità di insediamento in città dipende da vari fattori fra cui l’esistenza di vegetazione,disponibilità di acqua, suolo libero. Sono inoltre necessarie socialità, abitudini onnivore ed esplorative, familiarità con gli esseri umani. La città, come ambiente ecologico, è in grado di offrire al colombo urbano, più che ad ogni altra specie, un surrogato del suo habitat naturale ancestrale.
Le costruzioni urbane sono ricche di anfratti, che rappresentano un ottimo rifugio dove i piccioni possono trovare infinite possibilità di nidificazione. La città offre condizioni microclimatiche favorevoli e grandi disponibilità di cibo e ripari che inducono una perdita di contatto con l'ambiente naturale e rurale. Alti numeri di colombi si addensano negli edifici in cattivo stato di manutenzione o abbandonati. In questi luoghi i colombi costituiscono vere e proprie colonie, moltiplicandosi e costruendo i nidi con penne, feci e materiali vari (erbe, fuscelli, plastica e tessuti).
1.6 ALLE VAMENTO
Le razze attualmente allevate, derivano dalla domesticazione del piccione selvatico avvenuta nei secoli passati; si distinguono da questo per la maggiore variabilità del mantello e per la sua mansuetudine ed adattabilità alla vita con l'uomo.
È una specie capace di orientarsi egregiamente per ritrovare la sua colombaia, caratteristica che è stata fruttata e accentuata con la selezione ad opera degli allevatori. Le capacità di volo di un piccione sono impressionanti: in condizioni di tempo ottimale può percorrere ininterrottamente anche 1500 km ad una media di 60 km/h per ritornare alla colombaia di origine a cui rimane legato per tutta la vita (Ravazzi, 1995) .
I piccioni vengono allevati per tre scopi principali: la trasmissione di messaggi, per la produzione di carne ed a scopo ornamentale.
Nell'antichità i piccioni venivano costantemente utilizzati per trasportare messaggi in assenza di tecnologie alternative. L'importanza di questi animali è diminuita con l'utilizzo delle moderne tecniche di comunicazione, tuttavia l'uomo non ha mai smesso di allevarli sia per passione sia per lo studio delle loro capacità di orientamento.
L’allevamento del colombo leggero è sempre stato fatto in colombaie, lasciandolo libero di volare per i campi alla ricerca di cibo, somministrandogli solo una piccola quantità di granaglie. Per le razze pesanti e ornamentali, poco inclini al volo, l’allevamento è analogo a quello del pollame.
Attualmente esistono allevamenti intensivi o semi intensivi dove i piccioni sono tenuti in colombaie chiuse o in voliere, alimentati unicamente dall’uomo. Anche la riproduzione è controllata dagli allevatori, che creano le coppie, seguendole nella deposizione, cova e svezzamento dei nati.
Le razze allevate sono numerosissime ed ancora maggiori sono le varietà che si differenziano soprattutto per le caratteristiche del piumaggio, della livrea,dalla lunghezza, forma direzione delle penne, dalla conformazione di becco e testa e dal peso.
La maggior parte delle razze di colombi sono oggi raccolte in uno “Standards”, suddiviso in diverse categorie. Per quello che riguarda lo “Standards Italiano”, esso si suddivide in: colombi “Italiani”, di “Forma”, “Caruncolati”, “Gallina”, “Gozzuti”, di “Colore”, “Tamburi”, di “Struttura”, “Cravattati”, “Altovolanti e Capitombolanti”.
2. APPARATO URINARIO DEGLI UCCELLI
L’apparato urinario è costituito dai reni e dalle vie urinarie. Gli uccelli, a differenza dei mammiferi non sono dotati di vescica urinaria, ma di una cloaca dove si raccolgono anche materiali provenienti da apparato riproduttore e digerente.
Il rene è un organo emuntore, che interviene in modo determinante nell’omeostasi di liquidi interni e nell’eliminazione di prodotti terminali del catabolismo.
2.1 IL RENE
2.1.1 Embriogenesi
L’apparato urinario deriva dal mesoderma intermedio, che costituisce da ciascun lato della regione dosale dell’embrione, un cordone nefrogeno che, aumentando di volume, finisce per sporgere nella cavità celomatica.
I cordoni nefrogeni si estendono dalla regione cervicale alla sacrale e da questi originano in successione di tempo e di spazio tre sistemi emuntori: il pronefro, il mesonefro e il metanefro.
Il pronefro dei mammiferi rimane allo stato rudimentale,mentre negli uccelli evolve fino a raggiungere un certo grado di funzionalità.
Negli uccelli i primi tubuli pronefritici appaiono verso la 36° ora di incubazione e sono quasi totalmente regrediti verso la 55° ora, quando vengono sostituiti dal mesonefro, che rimane funzionante fino alla schiusa, per tutto il periodo embrionale (Veggetti, 1987). Le scorie cataboliche da esso filtrate, attraverso il dotto mesonefridico, raggiungono la cloaca e da qui si raccolgono nell’allantoide.
Il mesonefro viene poi sostituito progressivamente dal metanefro, o rene definitivo. Il metanefro è il terzo ed ultimo organo emuntore e rappresenta il rene funzionante nella vita post natale di rettili, uccelli e mammiferi.
2.1.2 Anatomia
Gli uccelli possiedono due reni appiattiti e allungati, che sono alloggiati nella cavità addominale in posizione retroperitoneale e accolti nelle docce tra le vertebre lombosacrali ed il sinsacro; tale spazio è definito depressione renale.
Il rene ha colore bruno, nei gallinacei è lungo 6-7 cm, nei piccioni 3-4 cm, ed appare diviso in tre porzioni di uguale lunghezza da due scissure di varia profondità. L’estremità craniale dei reni si trova in vicinanza dei polmoni, mentre quella caudale giunge fino al margine posteriore del sinsacro.
La superficie ventrale è percorsa da solchi dove sono alloggiate le vene iliache esterne, la vena renale caudale, la vena renale portale e la prima porzione dell’uretere. Entrambi i reni sono attraversati dal nervo ischiatico e da alcuni nervi spinali.
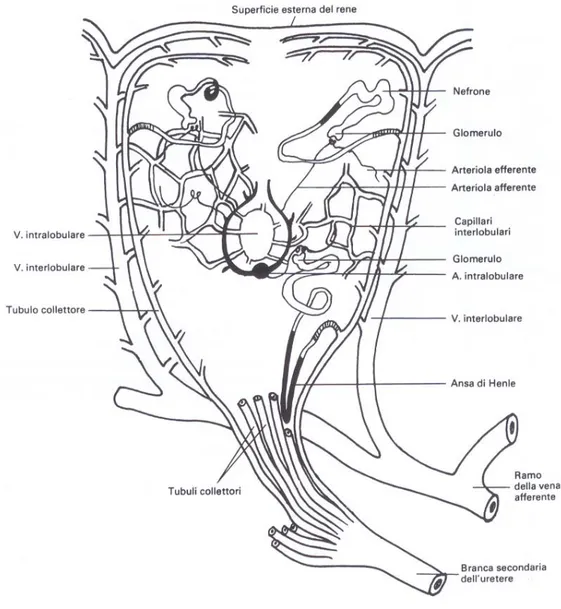
Ciascun rene è diviso in tre parti, craniale, media e caudale, ognuna delle quali contiene diversi lobi funzionali, ciascuno dei quali diviso il lobuli. Il parenchima è formato da lobuli piriformi, un certo numero di essi è superficiale, mentre la maggior parte hanno posizione profonda.
Osservando la superficie esterna dei reni è possibile apprezzare un disegno mammellonato, dato dalla parte apicale dei lobuli più superficiali. I lobuli sono delimitati da tessuto connettivale, dai rami terminali interlobulari della vena porta renale e dai tubuli collettori La disposizione dei nefroni e dei tubuli collettori nel lobulo è peculiare e può essere meglio compresa se si segue il destino delle vene interlobulari.
Da queste originano una miriade di capillari venosi sinusoidali che, a raggiera, si portano al centro del lobulo per aprirsi nella vena intralobulare, primo tratto della via efferente venosa. Negli spazi delimitati dalla rete venosa vengono a collocarsi i nefroni, anche loro con andamento a raggiera con i corpuscoli posti a mezza via tra la parte periferica e centrale del lobulo, i tubuli contorti prossimali, nelle porzioni periferiche, mentre quelli distali verso la zona centrale. I tubuli collettori periferici, confluiscono verso l’apice del lobulo unendosi a formare il fascio midollare, mentre la restante parte, formata da nefroni, rappresenta la corticale.
Le porzioni midollari di lobuli vicini si possono associare a formare un fascio di un numero limitato di dotti che sbocca in una diramazione dell’uretere.
La parte di rene drenata da una branca di uretere viene detta lobo.
Vascolarizzazione del rene:
La porzione craniale del rene è vascolarizzata dall’arteria renale craniale derivante da una branca dell’aorta, mentre le porzioni centrale e caudale sono supportate dalle arterie renali media e caudale che originano dall’arteria ischiatica.
Queste arterie una volta penetrate nel parenchima, si suddividono prima in rami extralobulari, dai quali originano le arterie intralobulari che decorrono al centro dei lobuli, in vicinanza della vena centrolobulare, originando numerose arteriole glomerulari afferenti che vascolarizzano i glomeruli.
Le arteriole glomerulari afferenti formano una rete capillare, la rete mirabile del glomerulo, sede dell’ultrafiltrazione del sangue.
Figura 1.2: Rappresentazione schematica dei principali vasi sanguigni destinati al rene (Da Botte‐Pelagalli 1982)
Dalla rete mirabile emergono le arteriole glomerulari efferenti che si ramificano lungo i tubuli costituendo la rete capillare intralobulare, dalla quale originano poi le vene efferenti intralobulari che si trovano al centro del lobulo e decorrono in senso inverso alle arterie. In prossimità della midollare alcune arteriole efferenti diventano rette spingendosi tra i dotti collettori.
Nei vertebrati inferiori la circolazione renale è ulteriormente complicata dalla presenza del
sistema portale renale. La funzione del sistema portale, presente in questi animali e
assente nei mammiferi, sembra sia quella di facilitare l’eliminazione dei cataboliti azotati a livello dei tubuli contorti.
La prima reale testimonianza della funzione del sistema portale renale è stata fornita da Sperber, che ha dimostrato che il rosso fenolo iniettato nella vena iliaca esterna di un arto, viene rilevato nell’uretere dello stesso lato dell’iniezione, prima che nell’uretere del lato opposto (Sperber, 1948). Questo dimostra la presenza di un flusso diretto di sangue dalle vene iliache al rene dello stesso lato.
Il sistema portale è costituito da una vena porta craniale ed una caudale. La vena porta caudale origina dall’unione delle vene iliache interna ed esterna, ischiatica e mesenterica caudale che raccolgono il sangue venoso refluo da coda, arti, pelvi e porzione caudale di intestino e rachide. La vena porta craniale origina invece da una branca della vena ischiatica esterna e si unisce alla porta caudale per formare un anello venoso localizzato ventralmente al rene.
Da questo anello vascolare si dipartono vasi che danno luogo alle vene renali afferenti, dalle quali originano le vene interlobulari, che si continuano nei capillari intralobulari, avvolti intorno ai tubuli contorti.
Un rapporto anatomico così intimo fa sospettare l’utilizzo preferenziale di questa via per eliminare alcune sostanze con la secrezione.
Ricerche sperimentali hanno evidenziato che prodotti, come l’acido urico, che sono eliminati soprattutto per secrezione lungo il tubulo, se inoculati in un arto si ritrovano nell’urina raccolta dal rene omolaterale (Botte-Pelagalli, 1982).
Il sangue portato da questo sistema può attraversare il rene, ma esistono dei meccanismi in grado di impedire questo percorso, attraverso connessioni dirette tra le vene portali e la cava caudale.
Esiste infatti una valvola portale renale localizzata tra le iliache comuni a regolare il flusso ematico venoso al rene.
La valvola portale renale è unica nel suo genere, in quanto è l’unica struttura intravascolare presente nei vertebrati a possedere una muscolatura liscia ed un supporto nervoso (Akester- Mann, 1969).
Quando la valvola è chiusa la maggior parte del sangue venoso passa attraverso il rene, questo accade quando il soggetto è a riposo. Quando la valvola è aperta il sangue venoso confluisce nella vena cava caudale bypassando il rene. Il sangue proveniente dall’anello venoso può saltare il filtro renale fluendo nella vena mesenterica caudale o nel seno venoso vertebrale interno.
Le valvole destra e sinistra del sistema portale renale possono operare in maniera indipendente ( Lierz, 2003).
È stato dimostrato che la valvola riceve una importante innervazione sia da fibre adrenergiche che colinergiche. Entrambi i tipi di fibre sono distribuiti uniformemente all’interno della valvola. Un confronto fra l’innervazione dei tessuti valvolari e la parete adiacente della vena, mostra che la valvola possiede una notevole innervazione rispetto alla vena (Akester- Mann, 1969).
La stimolazione adrenergica provoca il rilassamento dei muscoli valvolari, mentre quella colinergica causa contrazione. La muscolatura della vena porta renale si contrae sotto stimolazione adrenergica, regolando così il flusso ematico.
Sotto stimolazione simpatica la valvola è aperta ed il sangue bypassa il rene; mentre sotto stimolo parasimpatico aumenta la resistenza valvolare ed il sangue entra nel sistema portale renale (Lierz, 2003).
2.1.3 Istologia
Il nefrone:
Il nefrone rappresenta l’unità funzionale del rene ed è costituito dal corpuscolo di Malpighi, i tubuli contorti prossimale e distale e il dotto collettore. Negli uccelli il tratto intermedio, corrispondente all’ansa di Henle dei mammiferi, ha sviluppo vario. Nel rene infatti si individuano due tipi di nefroni: quelli di tipo corticale e quelli di tipo midollare.
I nefroni di tipo corticale sono numerosi, si dispongono nella zona più lontana dalla midollare e presentano una parte intermedia poco sviluppata o assente; sono detti anche
nefroni di tipo rettiliano.
I nefroni midollari che prendono il nome dalla loro disposizione in vicinanza della midollare, sono meno numerosi, possiedono glomeruli voluminosi e una parte intermedia ben sviluppata a formare un’ansa che si insinua tra i tubuli collettori della midollare, questi nefroni somigliano molto a quelli dei mammiferi.
Facendo una sezione del lobulo interessante corticale e midollare, i glomeruli appaiono distribuiti a ferro di cavallo intorno alla vena centrale con quelli di maggiori dimensioni in vicinanza della midollare.
Il corpuscolo di Malpighi, , è costituito dal glomerulo e dalla capsula di Bowman. Il glomerulo è formato da un ciuffo di capillari, intrecciati fra loro, ricoperti da epitelio che costituisce la lamina viscerale della capsula di Bowman. L’epitelio che riveste i capillari si ripiega a costituire la lamina parietale; fra capillari e parete si delimita così la spazio capsulare o spazio di Bowman che raccoglie l’ultrafiltrato del sangue. Questa struttura è analoga a quella dei mammiferi, ma il corpuscolo è meno ricco di capillari e quindi possiede minor area di filtrazione ( Botte-Pelagalli, 1982).
La lamina viscerale aderisce alle anse capillari, dalle quali è separata mediante una spessa membrana basale. Le sue cellule sono connesse a quelle della lamina parietale in corrispondenza del polo vascolare.
Al centro del glomerulo sono presenti numerose cellule mesangiali, spesso connesse a quelle endoteliali e separate da strutture connettivali simili a quelle della membrana basale ( Botte-Pelagalli, 1982).
Alcuni autori ritengono che queste cellule abbiano una funzione di sostegno in quanto secernerebbero un materiale, che ricopre l’endotelio, simile alla lamina basale; altri invece sostengono che le cellule mesangiali possono esplicare attività fagocitaria e rimuovere frammenti di lamina basale usurata impedendo che questa diventi eccessivamente spessa ( Dellmann-Brown, 1981).
Gli elementi epiteliali viscerali della capsula di Bowman, chiamati podociti, hanno forma cuboidale, con nucleo rotondo, citoplasma ricco di reticolo endoplasmatico rugoso e mitocondri. Possiedono dei prolungamenti citoplasmatici, i pedicelli, applicati alla
membrana basale senza però rivestirla completamente, restano quindi degli spazi liberi attraverso i quali possono passare le sostanze.
La membrana basale è formata da tre strati con il centrale più denso caratterizzato da materiale granulare e fibrillare, una lamina esterna a contatto con i prolungamenti dei podociti ed infine una lamina interna posta fra la lamina densa e le cellule endoteliali (Pak Poy-Robertson, 1957; Banks, 1991) .
L’endotelio capillare possiede cellule sottili ed espanse che aderiscono intimamente alla membrana basale. In alcuni punti sono presenti fenestrature di 400-1000 Å di diametro. L’epitelio del foglietto parietale è di tipo pavimentoso semplice, che diventa cubico al polo urinifero.
Il tubulo contorto prossimale ha un decorso tortuoso e una discreta lunghezza,ha un diametro di 35-45µ e può essere distinto in una regione prossimale e una distale, la quale possiede pareti di maggior spessore.
La membrana basale è facilmente distinguibile e colorabile con colorazioni elettive per polisaccaridi. L’epitelio è costituito da cellule cilindriche, con nucleo basale rotondeggiante provvisto di uno o due nucleoli e citoplasma ricco di mitocondri ed organuli, soprattutto lisosomi. A livello del margine libero la membrana cellulare costituisce un orletto a spazzola.
L’osservazione al microscopio elettronico di questa porzione ha svelato che tale orletto è costituito da numerosi microvilli, il citoplasma sottostante è ricco di vacuoli e tubuli inframezzati da invaginazioni della membrana cellulare. Delle invaginazioni analoghe sono presenti anche in vicinanza della lamina basale e della porzione più profonda di contatto fra cellule adiacenti; nelle restanti zone di contatto invece, gli elementi sono uniti da apparati giunzionali (Botte-Pelagalli, 1982).
Nella regione terminale del tubulo contorto prossimale, le cellule sono meno ricche di mitocondri. Questa porzione nei nefroni ad ansa lunga si assottiglia progressivamente e si approfonda nella midollare.
Il tratto intermedio o ansa di Henle, presente solo nei nefroni di tipo midollare, comprende una parte sottile ed una spessa, che si continua nel tubulo contorto distale. Il tratto sottile è molto breve ed è rivestito da epitelio cubico ciliato; mentre il tratto spesso si distingue per le cellule più voluminose e rotondeggianti prive di ciglia. Il tubulo contorto distale è rivestito di cellule con nucleo rotondeggiane e ricche di mitocondri, che tendono a
protrudere nel lume. La membrana cellulare possiede invaginazioni basali e laterali, ma a livello apicale ha pochi microvilli e vescicole di micropinocitosi.
Nel nefrone degli uccelli è presente l’apparato iuxtaglomerulare, costituito dalla macula densa che a livello del polo vascolare del glomerulo si associa alle cellule del mesangio. La macula densa è una porzione della parte iniziale del tubulo contorto distale, caratterizzata da cellule epiteliali alte e ricche di nuclei. La parte terminale del tubulo contorto distale si raccorda mediante un tubulo di connessione al sistema dei dotti collettori. Le cellule di questo tubulo presentano nella porzione apicale delle gocce di mucopolisaccaridi acidi e la loro membrana è povera di interdigitazioni. Il lume è irregolare con tracce di secreto mucoso.
I dotti collettori:
Trasportano l’urina dal nefrone all’uretere e probabilmente ne favoriscono la concentrazione attraverso il riassorbimento di acqua. Si distinguono i dotti interlobulari e quelli midollari. Gli interlobulari sono 12-20 per lobulo, si dirigono verso la regione midollare raccogliendo i vari tubuli di connessione ed anastomizzandosi tra loro. Sono rivestiti di epitelio con cellule cubico-cilindriche contenenti granuli mucinici nella porzione apicale. I dotti collettori midollari originano da quelli interlobulari, sono di maggiori dimensioni e possiedono cellule epiteliali ben distinte e infarcite di mucine, con nucleo basale rotondeggiante. Le mucine presenti nei dotti collettori servono probabilmente a lubrificare il lume in modo da impedire la stasi e la precipitazione degli urati.
2.2 URETERI
Da ciascun rene origina un uretere diviso in una parte renale e in una pelvica. La prima parte dell’uretere decorre sulla superficie ventrale del rene e riceve lo sbocco di branche collaterali connesse alla midollare dei singoli lobuli renali.
La parte pelvica è rappresenta il prolungamento di quella renale nel momento in cui si distacca dal margine posteriore del rene.
L’uretere si porta caudalmente fino a sboccare in cloaca, sulla volta dell’urodeo; da qui l’urina per giungere all’esterno deve attraversare il proctodeo.
Gli ureteri possiedono una muscolatura liscia che consente, per mezzo di movimenti peristaltici, la progressione dell’acido urico verso l’urodeo.
2.3 CLOACA
La cloaca è un organo impari, mediano diviso un maniera poco netta in tre cavità: coprodeo, urodeo e proctodeo.
La cloaca è in rapporto con la parete addominale caudale, con le anse intestinali e con la volta della cavità addominale.
Il coprodeo è il distretto più craniale, continua caudalmente il retto dal quale è separato per mezzo di una plica mucosa. Il coprodeo è rivestito di mucosa provvista di villi, simile a quella del retto.
L’urodeo è posto fra coprodeo e proctodeo dai quali è diviso da due pliche trasversali. La plica anteriore è abbastanza sviluppata e può comportarsi come un diaframma regolando la comunicazione con il coprodeo.
L’urodeo presenta sulla parete dorsale le aperture degli ureteri e lateralmente a queste, nel maschio, lo sbocco dei deferenti. Nella femmina sulla parete laterale sinistra si trova l’apertura dell’ovidutto sinistro, l’unico sviluppato nella maggior parte degli uccelli. La mucosa è simile a quella del coprodeo,data la medesima origine embriologica dall’endoderma.
Il proctodeo rappresenta la parte terminale della cloaca, sulla sua superficie dorsale accoglie lo sbocco della borsa di Fabrizio, un diverticolo che nell’animale giovane funge da organo linfopoietico. Il proctodeo è rivestito da mucosa con epitelio cilindrico semplice e origina dall’ectoderma; termina a livello dell’orificio cloacale esterno.
2.4 DIFFERENZE DI SPECIE
Nell’anatra e nell’oca la divisione craniale è sottile, mentre quella caudale è lunga ed espansa. I reni sono lunghi 9 cm circa nell’anatra e 11cm nell’oca. L’uretere emerge dalla superficie ventrale dell’organo piuttosto caudalmente.
Nel pappagallo i due reni sono spesso fusi nella divisione posteriore dove inglobano l’aorta. Gli ureteri sono nascosti alla loro origine, dalle vene renali.
2.5 FISIOLOGIA
Gli uccelli hanno un metabolismo molto intenso, perciò i cataboliti devono essere eliminati rapidamente; per questo motivo dispongono di reni proporzionalmente più voluminosi di quelli di mammiferi e rettili (1- 2,6% del peso totale) (Croce, 2002).
Gli uccelli, come i rettili, sono animali uricotelici, cioè a differenza dei mammiferi, l’azoto viene escreto per il 60-70%, sotto forma di urati, il 5-10% come urea (Aguggini-Beghelli, 2000) e la restante parte sotto forma di creatinina, aminoacidi e prodotti indeterminati (Botte-Pelagalli, 1982).
L’acido urico, che viene sintetizzato nel fegato è una peculiarità degli ovipari.
L’uovo infatti, scambia solo gas con l’ambiente esterno, mentre gli altri cataboliti devono essere accumulati nell’allantoide che è in grado di trattenere gli urati insolubili impedendone la diffusione all’uovo. Gli urati consentono all’embrione di riutilizzare l’acqua delle urine in quantità molto maggiore che non nel caso fosse eliminata urea. Probabilmente la selezione ha mantenuto questa caratteristica anche nell’adulto allo scopo di economizzare l’acqua (Croce, 2002).
L’acido urico, principale composto azotato presente nell’urina degli uccelli, viene
sintetizzato principalmente dal fegato per ossidazione dell’ipoxantina a xantina e quindi ad acido urico. La xantina deidrogenasi, enzima che promuove la conversione della xantina ad acido urico, è presente in minima parte anche nel rene che potrebbe quindi concorrere alla formazione del catabolita. Nel tubulo renale l’acido urico è eliminato essenzialmente per secrezione e si ritrova sotto forma di urati al pH fisiologico del sangue. Lungo il nefrone la concentrazione degli urati aumenta sia per la secrezione, sia per il riassorbimento di acqua nel tubulo prossimale; nonostante ciò gli urati non tendono a precipitare. La spiegazione di ciò deriva dal fatto che l’acido urico escreto dagli uccelli assume la forma di piccole sfere contenenti proteine e ioni inorganici (Braun-Pacelli, 1991; Casotti-Braun, 2004)
La concentrazione di urati può così salire ad oltre il 2% ed è mantenuta stabile dalla presenza delle mucine presenti nelle vie urinarie.
Le sfere sono composte da 65% di acido urico e 5% da varie proteine di cui la più rappresentata è la siero albumina (Braun-Pacelli, 1991). Studi effettuati con il microscopio a fluorescenza evidenziano la distribuzione in anelli concentrici delle proteine all’interno delle sfere urinarie (Casotti-Braun, 2004).
La concentrazione plasmatica di urati resta costante fino a che l’animale non è disidratato o i tubuli renali sono pieni di cristalli di urati. Gli urati continuano ad essere escreti nei tubuli, ma, in tal caso, non lasciano il rene per mancanza di flusso ureterico (Lierz, 2003). Il colore dell’acido urico, in uccelli sani, va dal bianco puro, al crema, fino al beige chiaro. I cambiamenti di colore più significativi si hanno in corso di patologie epatiche nelle quali il colore vira al giallo sporco o al verde pisello ( Don Harris, 1991).
L’urea presente nelle urine, non dà origine all’acido urico, ma sembra derivi principalmente dal metabolismo dell’arginina sia a livello epatico che renale, viene eliminata soprattutto per filtrazione.
I livelli di creatinina plasmatica negli uccelli sono generalmente bassi, perché la creatina viene in gran parte escreta prima della conversione in creatinina. La creatinina viene escreta con la filtrazione glomerulare e riassorbita a livello tubulare, in questo modo la concentrazione plasmatica si mantiene costante (Lierz, 2003).
La presenza di aminoacidi dipende dal tenore proteico della dieta, ma in minima parte sono sempre dosabili.
Il rene interviene attivamente per conservare costante il pH del sangue regolando l’escrezione di ioni formanti acidi o basi. L’urina può presentare un oscillazione di pH da 5 a 8, ma è sempre meno acida di quanto dovrebbe essere in rapporto all’eliminazione di acido urico, perché questo è presente sotto forma di sali poco dissociati.
Il potassio è secreto dal tubulo distale quando l’urina è acida (pH 5); in questo caso viene anche prodotta ammoniaca che viene escreta come sali di ammonio.
Una delle funzioni principali del rene è quella di mantenere l’omeostasi, regolando l’eliminazione dell’acqua e degli ioni. In presenza di eccessi di acqua si ha urina ipotonica, mentre in sua carenza l’urina è ipertonica.
L’osmoregolazione negli uccelli è garantita dall’attività di rene, intestino, ghiandola del sale ed infine da pelle e apparato respiratorio.
I meccanismi che intervengono nella regolazione dell’escrezione dell’acqua si basano su una variazione della filtrazione glomerulare e sull’intervento dell’ormone antidiuretico ipofisario.
Negli uccelli l’arginina-vasotocina è più attiva della vasopressina nel favorire il riassorbimento di acqua nel tubulo contorto distale.
Negli uccelli la diuresi può essere indotta dalla somministrazione di acqua, soluzione fisiologica, glucosio o mannitolo.
La maggior parte degli uccelli producono urine solo modestamente iperosmotiche, sebbene le scorie azotate siano eliminate principalmente come acido urico e suoi sali. Questo perché nel rene gli urati, che hanno bassa solubilità, si combinano a formare le sfere urinarie in sospensione colloidale supersatura ed in questo modo non contribuiscono significativamente all’osmolalità delle urine ureteriche (Braun, 2003).
Ciononostante, alcune specie producono urina molto concentrata, fino quasi 6000 mOsm nel caso del passero delle praterie (Passerculus sandwichensis) (Sherwood-Klandorf-Yancey, 2006)
Nei casi di grave disidratazione il riassorbimento di acqua da parte del rene è completato dall’attivazione di un meccanismo fisiologico che causa il reflusso di urina nella cloaca e nel retto, sede dell’assorbimento dell’acqua, per cui riducendo la quota persa favorisce il ripristino dell’equilibrio idrico.
Quando l’urina entra in cloaca l’acido urico si combina con gli ioni presenti e precipita a formare una massa densa e biancastra contenente sodio, potassio, sali d’ammonio dell’acido urico e altri ioni. L’urina diventa così meno concentrata e libera acqua che può essere riassorbita a livello cloacale insieme al sodio attraverso meccanismi di trasporto attivo.
Clearance renale:
Come nei mammiferi i nefroni operano utilizzando meccanismi di filtrazione glomerulare, riassorbimento selettivo e secrezione.
Individualmente i nefroni degli uccelli hanno bassa velocità di filtrazione glomerulare, ma questi animali possiedono nel complesso un maggior numero di nefroni rispetto ai mammiferi, così la GFR complessiva risulta essere simile.
Il numero totale dei glomeruli varia da 30.000 di un passeriforme a 274.000-353.000 dei piccioni e 1.989.000 delle anatre, quindi da 4 a 15 volte più numerosi che nei mammiferi (Croce, 2002).
Il rene di un uccello di 100 g filtra approssimativamente, 11 volte l’intero contenuto idrico corporeo, ogni giorno; riassorbendo poi molta dell’acqua filtrata.
La GFR (velocità di filtrazione glomerulare) diminuisce con la disidratazione e con la somministrazione di arginina vasotocina, che negli uccelli rappresenta l’analogo dell’ormone antidiuretico dei mammiferi (Harr, 2002).
La filtrazione glomerulare è stata valutata utilizzando sostanze organiche, come l’inulina, che passano nelle urine senza essere riassorbite né secrete lungo il tubulo. Il valore di eliminazione dell’inulina è di 2 ml/min/Kg ed è considerato come indice di massima filtrazione di un composto. Sostanze con valori maggiori vengono escluse anche per secrezione, mentre nel caso inverso sono in parte riassorbite ( Botte-Pelagalli, 1982).
Il riassorbimento selettivo gioca un ruolo fondamentale nella funzione renale, esso varia molto a seconda dei prodotti, per cui si passa dal glucosio, che viene completamente riassorbito, ad una velocità di 5 mg/min/Kg; all’acqua che viene per il 99% riassorbita, fino a sostanze che passano totalmente nelle urine.
La secrezione è particolarmente importante per alcuni cataboliti, come l’acido urico che viene eliminato per il 93% per questa via.
Gli uccelli hanno la capacità di ridurre la filtrazione glomerulare, quando l’apporto idrico è insufficiente, allo scopo di ridurre la quota idrica persa con le urine.
L’abilità a concentrare le urine varia in maniera inversamente proporzionale alla taglia, così uccelli più piccoli hanno maggiore abilità a concentrare le urine
(Harr, 2002).
Il riassorbimento avviene a livello dei tubuli prossimali per opera di cellule con orlo a spazzola, ma presentando pochi mitocondri dovrebbero espletare una bassa attività e scarso riassorbimento. In questa specie la secrezione tubulare è un processo attivo che viene espletato con una intensità maggiore rispetto ai mammiferi.
3. PATOLOGIE RENALI
3.1
PATOGENESI
3.1.1 Cause congenite
Anche se non sono molto frequenti, le lesioni renali congenite possono essere occasionalmente segnalate negli uccelli.
La totale assenza dei reni è riportata, ma porta a morte embrionale precoce. Al contrario si può trovare con maggiore frequenza l’ipoplasia, con ipertrofia compensatoria della parte intatta (Tudor, 1979). Segni clinici si hanno solo se gran parte del rene è non funzionante e quindi la porzione ipertrofica non riesce a compensare la perdita di funzionalità (Lierz, 2003). Per questo motivo è spesso trovata casualmente durante le necroscopie e la porzione del rene che manca più frequentemente è quella craniale (Schmidt, 2006). E’ necessario differenziare l’ipoplasia congenita dall’atrofia ed ipotrofia: reni ipotrofici possono essere trovati in uccelli con ascite, o con altre patologie che provocano compressione sul parenchima renale. La diagnosi di aplasia o di ipoplasia evidenziate con endoscopia o urografia deve essere confermata dall’istologia (Lierz, 2003).
3.1.2 Cause nutrizionali e metaboliche
Una dieta con eccesso di calcio, proteine o vitamina D, oppure carente di vitamina A, può condurre a nefropatie o altre patologie degenerative renali.
Ipovitaminosi A può causare metaplasia squamosa dei tubuli renali e dell’epitelio ureterico (Lierz, 2003). L’uretere può poi ostruirsi con cellule cheratinizzate ed esitare in idronefrosi (Styles-Phalen, 1998).
L’eccesso di vitamina D3 invece, promuove la mineralizzazione metastatica dei visceri,
rene compreso (Pollock, 2006).
La lipidosi renale può essere secondaria a diete troppo ricche di grassi o patologie epatiche croniche. Macroscopicamente il rene appare pallido e microscopicamente è visibile la presenza di lipidi nelle cellule dell’epitelio tubulare. Nei capillari glomerulari sono generalmente presenti macrofagi contenenti lipidi (Schmidt, 2006).
3.1.3 Cause infiammatorie
L’amiloidosi è frequentemente associata a stati infiammatori cronici, come sepsi, gotta, artriti ed enteriti (Pollock, 2006), ma anche pododermatiti, aspergillosi e tubercolosi aviare (Styles-Phalen, 1998).
La deposizione di sostanza amiloide nelle membrane basali di tubuli e glomeruli è stata riscontrata nelle anatre. In molti casi fa parte di una amiloidosi generalizzata, che causa un severo danno glomerulare che esita poi in sindrome nefrosica con possibilità di gotta (Lierz, 2003).
Macroscopicamente il rene è aumentato di volume, pallido e friabile. Istologicamente l’amiloide è eosinofila o amorfa e può depositarsi nella membrana basale di glomeruli o tubuli e nella parete delle arterie e arteriole renali (Schmidt, 2006).
3.1.4 Cause infettive
Batteriche:
Molti batteri possono causare infezioni renali secondarie a patologie sistemiche. Fra i patogeni più comuni abbiamo Staphilococcus spp., Streptococcus spp., E.coli, Salmonella spp. ed altre enterobacteriacae, Pasteurella spp., Micobacterium avium e Chlamydophila psittaci (Lierz, 2003).
Le nefriti batteriche sono generalmente secondarie a patologie sistemiche ed i microrganismi arrivano al rene per via ematogena attraverso l’arteria renale o il sistema portale renale, mentre le infezioni di tipo ascendente sono meno frequenti (Pollock, 2006). In entrambi i tipi di infezioni il rene può risultare megalico con grado variabile di necrosi, ma nelle forme ematogene le lesioni si localizzano inizialmente a livello del glomerulo. Le lesioni istologiche, indicative di nefriti batteriche, includono la dilatazione tubulare con presenza di cellule infiammatorie. Quando la nefrite cronicizza, si evidenziano anche necrosi tubulare, formazione di cisti, fibrosi interstiziale con infiltrazione di cellule mononucleate. La necrosi dell’epitelio tubulare è spesso rilevante, ma in alcuni casi di nefriti batteriche può essere minima o assente (Schmidt, 2006).
Virali:
Il virus della bronchite infettiva ha tropismo per le cellule renali, ma anche altri virus come herpesvirus, paramyxovirus e reovirus possono infettare il rene (Lierz, 2003).
In corso di infezioni da Adenovirus, le lesioni microscopiche sono generalmente minime e vanno da una moderata infiltrazione interstiziale di cellule mononucleate, fino a vacuolizzazioni e necrosi delle cellule dell’epitelio tubulare.
Alcune cellule epiteliali tubulari possiedono nuclei cariomegalici contenenti grossi corpi inclusi francamente eosinofili o basofili.
Le infezioni da Polyomavirus, possono essere acute e il virus può essere isolato dal rene. Anche in questo caso le cellule dell’epitelio tubulare possono presentare cariomegalia con corpi inclusi.
Paramyxovirus-1 nei piccioni può causare nefrite interstiziale linfoplasmocitaria e necrosi tubolare con granuli e cilindri ialini all’interno dei tubuli (Schmidt, 2006).
3.1.5 Cause parassitarie
Le coccidiosi possono provocare nefriti granulomatose in varie specie di uccelli, come galliformi, colombiformi e passeriformi. Tanaisia bragai, un trematode può provocare ostruzione dei dotti collettori (Lierz, 2003).
3.1.6 Cause traumatiche
I traumi diretti al rene sono rari negli uccelli perché l’organo è ben protetto dallo scheletro. Se si sviluppa un ematoma renale può comprimere i nervi spinali provocando paresi degli arti. Si può avere schiacciamento renale durante distocia (Pollock, 2006).
3.1.7 Cause tossiche
A causa della presenza del sistema portale epatico e renale il rene degli uccelli è spesso bersaglio di tossine provenienti dall’intestino (Pollock, 2006).
Le nefrotossine, come metalli pesanti, micotossine, pesticidi e alcuni farmaci, causano comunemente necrosi tubulare renale. I segni clinici che comunemente si riscontrano sono letargia, vomito, polidipsia e poliuria, ma in caso di insufficienza renale acuta ci può essere anuria (Lierz, 2003).
3.1.8 Cause degenerative
La nefrosi è una patologia degenerativa del rene in assenza di infiammazione. Si sviluppa in seguito all’esposizione a tossine o secondariamente a infezioni che non necessariamente determinano involuzione renale( Lierz, 2003).
3.1.9 Neoplasie
Le neoplasie del rene sono riportate in molti uccelli, ma sono particolarmente comuni nei pappagalli ondulati (Melopsittacus undulatus) (Styles-Phalen, 1998).
I tumori renali primari riportati negli uccelli sono adenomi, adenocarcinomi e nefroblastomi. Il più comune è l’adenocarcinoma che talvolta causa osteolisi e sclerosi dell’ileo e del sinsacro e può potenzialmente infiltrare i tessuti vicini (Pollock, 2006). In molti casi sono riportati letargia e zoppia di un arto. Le masse tumorali determinano compressione dei plessi nervosi fino alla paralisi. Se il tumore renale è craniale può essere difficile differenziarlo da una neoplasia gonadica. In caso di sospetto di neoplasia ecografia ed endoscopia rappresentano un utile supporto diagnostico, mentre la biopsia è indispensabile per fare una corretta diagnosi (Lierz, 2003).
3.1.10 Urolitiasi
Lo sviluppo di calcoli urinari è comunemente associato a stati di grave disidratazione, altre cause includono eccessi di calcio nella dieta e presenza di infezioni. La presenza di uroliti a livello renale determina una ipertrofia compensatoria del restante tessuto renale. I calcoli post renali sono responsabili di compressione e ostruzione dell’uretere (Pollock, 2006).
3.1.11 Gotta
La gotta è più un segno di un problema renale che una distinta patologia, poiché insorge ogni volta che la concentrazione di urato di sodio eccede la sua solubilità. Oltre alle patologie renali, anche le proteine presenti nella dieta possono causare iperuricemia.
In corso di disidratazione il rene è ancora in grado di secernere acido urico, ma il basso flusso di urine nei tubuli provoca precipitazione dei cristalli, che provocano ostruzione postrenale, seguita da anuria ed oliguria. Questa è conosciuta come gotta renale, mentre l’iperuricemia provoca gotta articolare o viscerale (Lierz, 2003).
3.2 DIAGNOSI
3.2.1 Analisi delle urine
Spesso le patologie renali non vengono riconosciute, poiché le urine vengono eliminate insieme alle feci, quindi la poliuria, che è un sintomo piuttosto frequente, viene normalmente confusa con la diarrea (Croce, 2002).
Alterazioni urinarie possono rappresentare un’indicazione precoce di patologia renale (Lierz, 2003). L’urina utilizzabile a scopo diagnostico dovrebbe essere priva di urati, questo non sempre è possibile in condizioni normali, ma risulta più facile nei soggetti poliurici (Lierz, 2003).
Metodi di raccolta:
L’urina negli uccelli può essere ottenuta in vario modo. Il metodo più semplice consiste nel raccoglierla direttamente dal pavimento del ricovero utilizzando un substrato plastico o una carta non assorbente (Pollock, 2006).
Altre tecniche comprendono la raccolta delle urine direttamente dall’uretere (Nechay-Del Carmen Lutherer, 1968; Wideman-Braun, 1982) o dall’urodeo previa incannulazione (Haselma et al., 1988).
All’infuori dell’urina raccolta direttamente incannulando l’uretere, è difficile raccogliere campioni puri senza contaminazione fecale (Styles-Phalen, 1998).
L’urina raccolta dopo l’escrezione è significativamente diversa rispetto a quella ureterica per la presenza di contaminanti e per l’assenza di frazioni che vengono riassorbite, per questo motivo ne va tenuto conto al momento dell’interpretazione dei risultati.
colore:
Nella valutazione del colore dell’urina è importante tener conto del metodo di raccolta, infatti pigmenti presenti nelle feci possono contaminare l’urina cloacale (Pollock, 2006). Il colore normale delle urine è bianco o giallo chiaro, ma può variare in base alla presenza e alla concentrazione dei pigmenti disciolti.
I pigmenti biliari, fanno assumere all’urina una colorazione verde smeraldo o nerastra, la biliverdina colora le urine di verde chiaro, mentre il sangue, emoglobina e mioglobina di
rosso (Pollock, 2006). La biliverdinuria presente in corso di patologie epatiche, può essere talvolta associata anche ad aspergillosi sistemica (Lierz, 2003)
Osmolalità e peso specifico:
Le urine degli uccelli hanno bassa osmolalità, valori elevati si riscontrano solo in uccelli che si sono adattati ai climi desertici.
Le urine raccolte con cannula dall’uretere hanno osmolalità più bassa di quelle escrete, che subiscono riassorbimento di acqua nell’intestino e nella cloaca.
Studi svolti sui piccioni dimostrano che una osmolalità di 450 mOsmol/Kg corrisponde alla normale capacità di concentrazione di un uccello sano. Tale valore aumenta dopo 24 ore di privazione di acqua sia nei piccioni sani che in quelli poliurici senza danni renali (Lierz, 2003).
Il peso specifico dell’urina degli uccelli oscilla tra 1.005 e 1.020 g/mL (Styles-Phalen, 1998).
pH:
Il pH delle urine degli uccelli oscilla tra 6.0 e 8.0 e può essere influenzato dalla dieta e dal materiale cloacale (Phalen et al, 1990; Pollock, 2006).
Per i piccioni l’intervallo di riferimento è 5.5-6.9 (Halsema et al, 1988).
Acido urico ed ammoniaca:
La concentrazione di acido urico può essere misurata nel sopranatante, mentre nel sedimento si hanno concentrazioni troppo elevate (Lierz, 2003).
Creatinina:
La creatinina è presente solo in ridotta quantità nell’urina degli uccelli, essendo eliminata sotto forma di creatina (Sturkie, 1965).
Proteine:
Le urine contengono quantitativi significativi di proteine, ma queste sono racchiuse nelle sfere urinarie. Dopo la centrifugazione e la rimozione del sedimento, nel sopranatante rimangono solo tracce di proteine (Styles-Phalen, 1998).
Concentrazioni di proteine nelle urine di 2 g/L sono considerati normali. La proteinuria è un segno di aumento della permeabilità glomerulare, spesso associata a glomerulonefriti (Lierz, 2003).
Il dosaggio nel piccione oscilla fra 0,11 e 1,99 g/L (Halsema et al, 1988).
Glucosio:
La concentrazione di glucosio nell’urina è generalmente bassa e non può essere rilevata con gli sticks urinari. Al contrario se gli sticks reagiscono positivamente viene fatta la diagnosi di glicosuria. Cause di glicosuria possono essere diabete mellito e pancreatite (Lierz, 2003).
Enzimi Urinari:
I tessuti renali contengono Lattato-Deidrogenasi, Glutammato-Deidrogenasi, Aspartato-Aminotrasferasi, Alanino-Aspartato-Aminotrasferasi, Creatina-Chinasi e Fosfatasi Alcalina, che vengono eliminati per via urinaria. Elevate concentrazioni di questi enzimi indicano un danno renale, ma non sono ancora disponibili gli intervalli di riferimento (Lierz, 2003).
Sedimento:
Il sedimento contiene normalmente molte cellule squamose epiteliali, urati, ossalato di calcio e cristalli di sulfonamide, un basso numero di leucociti e globuli rossi e pochi batteri probabilmente di origine fecale o cloacale (Pollock, 2006).
3.2.2 Analisi sangue
Variazioni dei valori plasmatici avvengono tardi in corso di patologie renali (Lierz, 2003). Per identificare sepsi e nefriti batteriche la coltura a partire dal sangue dell’animale è più indicata dell’urinocoltura (Pollock, 2006).
Acido urico ed urea:
L’acido urico e l’urea possono essere misurati, ma aumenti patologici di acido urico vengono rilevati solo dopo che il 70 % della funzionalità renale è stata persa, mentre l’urea può essere utilizzata solamente per svelare disidratazione (Lierz, 2003). Quindi nessuno di questi due parametri è in grado di rivelare una disfunzione renale, soprattutto se precoce.
Creatinina:
Il dosaggio della creatinina è di scarso valore diagnostico negli uccelli, perché in questi animali la maggior parte della creatina viene escreta prima della sua conversione in creatinina e quindi i suoi livelli plasmatici risultano sempre molto bassi (Lierz, 2003).
Elettroliti:
L’iperkaliemia può essere rilevata in corso di insufficienza renale acuta, ma i risultati per questo parametro possono essere influenzati dalla raccolta e gestione del campione ematico (Lierz, 2003).
L’ipernatriemia può essere osservata in corso di disidratazione o a seguito di eccesso di sodio con la dieta. Iponatriemia può indicare insufficienza renale(Lierz, 2003).
Il calcio viene normalmente riassorbito dopo la filtrazione glomerulare; in corso di insufficienza renale la perdita di calcio può essere ingente risultando in ipocalcemia (Lierz, 2003).
La riduzione del tasso di filtrazione glomerulare può condurre ad innalzamenti della concentrazione plasmatica del fosforo inorganico (Lierz, 2003).
3.2.3 Radiologia
Il rene degli uccelli è difficile da valutare radiograficamente a causa della sua posizione all’interno del sinsacro, in ogni caso la proiezione laterale risulta migliore della ventrodorsale, perché in quest’ultima l’intestino si sovrappone al rene. Il segno radiografico più evidente di nefromegalia è l’aumento di volume della divisione craniale (Pollock, 2006). L’urografia discendente può essere utilizzata per differenziare il rene e l’uretere da altre strutture o per svelare ostruzioni ureteriche (Lierz, 2003).
3.2.4 Ecografia
L’ecografia renale è generalmente impossibile da effettuare normalmente negli uccelli a causa della posizione dorsale dei reni e della presenza dei sacchi aerei. In uccelli di grandi dimensioni è stato tentato un approccio transcloacale (Pollock, 2006)
3.2.5 Endoscopia
L’endoscopia è molto utile per la diagnosi di patologie degli organi interni perché determina una diretta visualizzazione ed interpretazione delle lesioni. Possono essere valutati con questa tecnica la taglia, il colore, la forma e la superficie del rene. È possibile inoltre evidenziare un’ eventuale ostruzione ureterica e effettuare un biopsia valutando visivamente il rene. La presenza di piccole macchie bianche o gialle sulla superficie del rene sono indice di gotta o di disturbi dell’escrezione.
Un uretere ripieno e gonfio è indicativo di ostruzione o disidratazione (Lierz, 2003).
3.2.6 Biopsia renale
La biopsia renale è il miglior test diagnostico per svelare patologie renali. Tale tecnica risulta indicata in caso di poliuria/polidipsia, alterazioni della biochimica sierica, anomalie delle analisi delle urine, alterazioni radiografiche. Il sito standard di ingresso è il sacco aereo toracico caudale (Pollock, 2006).
Il sito bioptico spesso consigliato è la divisione craniale per le sue dimensioni e la buona visibilità (Müller et al., 2004). L’arteria craniale renale però decorre più superficialmente rispetto alle altre, quindi i rischio di lacerare l’arteria durante la biopsia risulta più alto (Pollock, 2006).
4. PARTE SPERIMENTALE
4.1 SCOPO DELLA TESI
Attualmente, per quanto riguarda gli uccelli, non sono disponibili metodi scarsamente invasivi e precoci per diagnosticare patologie renali; l’unica tecnica valida è quella della biopsia renale. Gli studi effettuati fino ad ora si concentrano soprattutto su polli o uccelli ornamentali. Il piccione rappresenta un modello di studio, data la sua grande distribuzione sia nelle aree urbanizzate che rurali,che quindi consente di reperire un congruo numero di soggetti di svariata provenienza. L’analisi delle urine con l’utilizzo di sticks è spesso risultata non idonea all’applicazione negli uccelli. Nei mammiferi per lo studio delle proteine urinarie viene da diverso tempo utilizzata la metodica SDS-PAGE, tale tecnica da precedenti studi è risultata applicabile anche alle urine di pollo (Janes-Braun, 1997). Nel nostro lavoro abbiamo confrontato l’SDS-PAGE delle urine e il referto istologico della biopsia renale. Lo scopo di questa tesi è la messa a punto di un protocollo diagnostico a partire dalle urine.
Gli obiettivi che questo lavoro si pone sono rappresentati da:
• Descrizione delle procedure sperimentali eseguite su ciascun animale
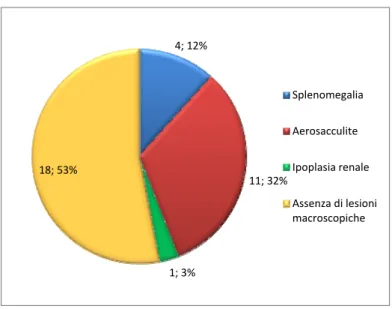
• Descrizione delle lesioni rilevate durante l’esame clinico dell’animale e calcolo della frequenza.
• Descrizione delle lesioni rilevate durante la celioscopia o necroscopia e calcolo della frequenza.
• Raccolta di urine ureteriche da animali morti.
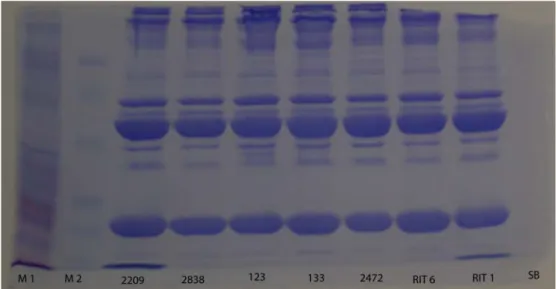
• Applicazione della metodica SDS-PAGE alle urine ed al plasma di piccione.
• Correlazione tra le proteine presenti nelle urine ureteriche e quelle direttamente escrete dall’animale.
• Correlazione tra l’uroprotidogramma e le proteine plasmatiche.
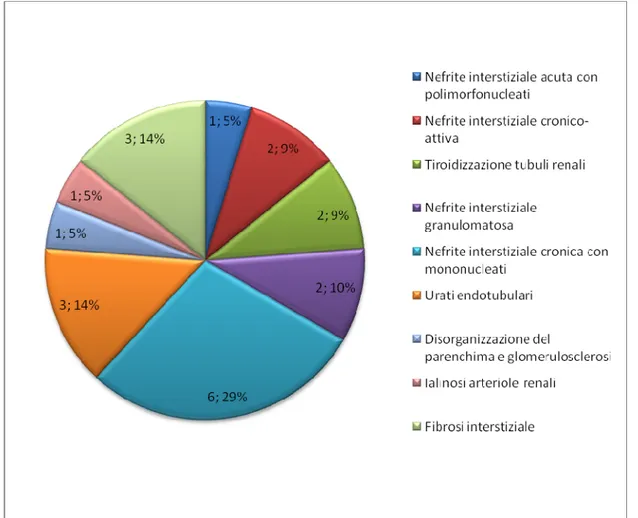
• Studio dell’incidenza dei vari tipi di lesioni riscontrate dall’analisi istologica del rene.
4.2 MATERIALI E METODI
Il lavoro di campionamento è stato svolto presso il CRUMA (Centro Recupero Uccelli Marini ed Acquatici) di Livorno. I dati sono stati raccolti nel periodo dicembre 2007 -febbraio 2009.
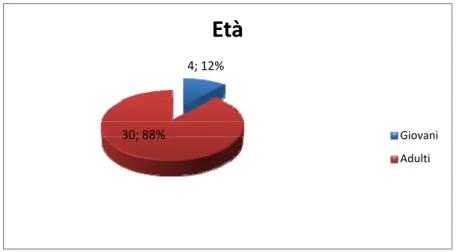
Lo studio è stato condotto su 34 piccioni (Columba livia), ricoverati presso il centro per svariati motivi. In certi casi si trattava di soggetti giovani, ma la maggior parte era rappresentata da soggetti adulti.
14 soggetti presentavano una sintomatologia che faceva ipotizzare un interessamento renale (poliuria-polidipsia, oliguria), mentre i restanti animali avevano subito traumi articolari con danni tali, da renderli incapaci di volare. Questi soggetti presentavano un apparato urinario apparentemente sano.
Lo scopo del centro di recupero è quello di curare gli uccelli in modo da poterli reintrodurre nel loro ambiente nel minor tempo possibile. Gli animali possono essere liberati solo dopo completa guarigione e dopo aver riacquistato la capacità di compiere le attività ed i movimenti tipici della specie. Per questo motivo i piccioni che avevano subito un trauma invalidante e che quindi non potevano essere nuovamente liberati, venivano sottoposti ad eutanasia.
I soggetti utilizzati risiedevano in voliere poste all’esterno o in stabulari all’interno dell’edificio.
Per facilitare le operazioni di restrizione alimentare e di campionamento, i piccioni venivano trasferiti in ricoveri individuali 24-48 ore prima dell’inizio dello studio.
Di tutti i soggetti campionati, i sintomatici sono stati sottoposti ad un protocollo che comprendeva:
• Campionamento delle urine escrete • Protocollo anestesiologico
• Prelievo ematico e gestione dei campioni • Celioscopia
Nei soggetti che dovevano essere sottoposti ad eutanasia, è stato applicato un protocollo alternativo che comprendeva:
• Campionamento delle urine escrete • Protocollo anestesiologico
• Prelievo ematico e gestione dei campioni • Eutanasia
• Necroscopia
• Campionamento di parenchima renale.
In due di questi soggetti durante la necroscopia, che avveniva immediatamente dopo l’eutanasia, sono state raccolte le urine ureteriche.
DESCRIZIONE DELLE FASI
• CAMPIONAMENTO URINE ESCRETE
Come noto dall’anatomia, gli uccelli convogliano le urine in cloaca, dove affluiscono anche le secrezioni ed escrezioni dell’apparato digerente e riproduttore.
Per ridurre la quota di materiale di provenienza intestinale i piccioni sono stati sottoposti a digiuno preventivo per almeno 24 ore, continuando comunque a somministrare acqua a volontà.
L’urina è stata raccolta dal pavimento dei ricoveri, costituito di materiale liscio ed impermeabile, utilizzando una siringa alla quale è stato tolto l’ago e posta in una eppendorf da 1 ml identificata con il codice attribuito a ciascun piccione.
Per un soggetto particolarmente poliurico sono stati raccolti tre campioni a distanza di un’ ora l’uno dall’altro, ottenendo quindi un totale di 36 campioni.
Dopo averne raccolto un quantitativo sufficiente, il materiale veniva centrifugato per 10 minuti a 10000 rpm. Il surnatante di ciascun campione veniva raccolto in una nuova eppendorf e congelato, mentre il sedimento veniva scartato perché non considerato utile ai fini della ricerca. Nel caso di campioni abbondanti era prevista la divisione in due aliquote che venivano destinate rispettivamente all’esame biochimico e all’SDS-PAGE.
• PROTOCOLLO ANESTESIOLOGICO
Dopo aver completato la raccolta delle urine gli animali venivano portati uno alla volta nella sala chirurgica presente nel centro.
Prima di poter effettuare le operazioni successive risultava necessaria la totale sedazione degli animali, effettuata tramite anestesia gassosa con isoflurano (2-3%, a seconda dell’efficacia dell’anestesia) ed ossigeno (1,5 L/min).
Negli uccelli infatti è consigliabile utilizzare come agente anestetico l’isoflurano associato ad ossigeno e protossido d’azoto o solo ossigeno, perché la sua rapida reversibilità e scarsa capacità di legarsi ai tessuti permette di avere un alto margine di sicurezza; in caso di arresto respiratorio infatti è sufficiente ventilare il paziente con ossigeno puro per riattivare la respirazione. L’alotano invece è da considerarsi pericoloso, sia per l’alta velocità con cui si lega ai tessuti, soprattutto adiposo ed epatico, sia per la scarsa reversibilità che risulta estremamente pericolosa in caso di arresto respiratorio (Croce, 2002).
La miscela di gas anestetico veniva somministrata attraverso una mascherina ricavata dalla parte prossimale di una bottiglietta da 0,5 l di plastica, la cui estremità più larga era parzialmente occlusa con un pezzo di guanto in lattice in modo da ridurre la possibilità di fuoriuscita dei gas nella sala operatoria.
Il collo della bottiglia veniva collegato alla macchina anestesiologica tramite un tubo corrugato dotato di un piccolo pallone di riserva ricavato da un profilattico.
La testa ed il collo dell’animale venivano introdotti nella mascherina e si attendeva la completa sedazione del soggetto.
• PRELIEVO EMATICO
Una volta anestetizzato l’animale, veniva effettuato un prelievo ematico a livello della vena giugulare.
Negli uccelli è possibile prelevare con sicurezza un campione pari all’1% del peso corporeo dell’animale (Murray,1997). Si rende comunque necessaria, una metodica
di prelievo adeguata, considerata anche la maggior fragilità e predisposizione alla lisi delle cellule ematiche aviarie rispetto a quelle dei mammiferi (Murray, 1997).
Negli Stati Uniti è pratica comune utilizzare per il prelievo un ago, contenente una piccola quantità di sodio eparina iniettabile al fine di ridurre la possibilità di coaguli.
Il prelievo comunque, se compiuto da mani esperte, non crea danni rilevanti alla parete vasale e consente di prelevare elevate quantità di sangue senza utilizzare eparina (Harr KE, 2002).
La vena giugulare destra è il sito di elezione per il prelievo, data la maggior dimensione di questo vaso rispetto al controlaterale. Con il piccione in decubito ventrale, veniva ruotata la testa verso destra ed il collo veniva afferrato da chi eseguiva il prelievo fra l’indice ed il medio della mano sinistra. Individuata la vena scostando con le dita le piume ed occludendo a valle il vaso, facendo una pressione con il dito pollice della mano sinistra alla base del collo, previa disinfezione con alcool, si procedeva al prelievo con la siringa tenuta nella mano destra.
Era utilizzato un ago da 27 G inastato su di una siringa da 1 ml per ridurre al minimo il rischio di ematomi. Una volta concluso il prelievo il dito utilizzato per occludere la vena veniva spostato sul sito di prelievo in modo da creare emostasi per compressione.
Prelevare il sangue con aghi di 25 gauge o di misura inferiore può causare emolisi da moderata a marcata, che potrebbe invalidare molte analisi biochimiche (Harr KE, 2002). Al fine del nostro utilizzo questo non creava problemi.
Talvolta la vena giugulare risultava difficilmente individuabile, in questi casi veniva utilizzato come sito di prelievo la vena alare che decorre a livello sottocutaneo nella faccia ventrale dell’articolazione omero radioulnare.
La regione veniva bagnata con alcool, in modo da visualizzare la vena sulla quale veniva effettuata una pressione nella regione a valle del sito di prelievo.
Veniva prelevato circa 1 ml di sangue, pari alla capacità massima della siringa utilizzata, che veniva trasferito, disinnestando l’ago dalla siringa, in una provetta da 1,3 ml contenente litio-eparina. Le provette erano contrassegnate con il codice attribuito al piccione, in modo da rendere i campioni facilmente identificabili. I campioni ematici venivano centrifugati a 4500 giri per 10 minuti ed il plasma così ottenuto trasferito in una eppendorf e congelato.
• EUTANASIA
L’eutanasia dei soggetti irrecuperabili è stata effettuata attraverso la somministrazione di Tanax® all’interno della vena alare.
• CAMPIONAMENTO DELLE URINE URETERICHE
La raccolta delle urine ureteriche è stata effettuata solamente in due dei soggetti sottoposti ad eutanasia, poiché si tratta di una pratica molto invasiva, con la tecnica da noi utilizzata, da non renderla applicabile in un soggetto in vita. La tecnica non è semplice per le ridotte dimensioni dell’uretere, inoltre il materiale raccolto nell’animale deceduto è molto esiguo, quindi sono stati necessari vari tentativi su altri cadaveri prima di riuscire ad ottenere i due campioni utilizzati nel lavoro.
Il campionamento delle urine ureteriche era comunque necessario per poter successivamente differenziare le proteine urinarie di pertinenza renale, da quelle che derivano da una contaminazione cloacale.
Per effettuare tale campionamento, il piccione veniva posizionato in decubito dorsale e veniva praticata una incisione a forma di “v” nella porzione bassa della cavità toracoaddominale.
Gli organi venivano rapidamente osservati per rilevare anormalità macroscopicamente osservabili.
Venivano quindi visualizzati gli ureteri sollevando il pacchetto intestinale. Gli ureteri venivano cateterizzati con un catetere intravenoso da 24 G ed il materiale in essi contenuto veniva aspirato raccordando al catetere una siringa. Le urine ureteriche così ottenute venivano processate con lo stesso metodo di quelle eliminate spontaneamente dall’animale e congelate.
• NECROSCOPIA E CAMPIONAMENTO DI TESSUTO RENALE
Nei soggetti deceduti veniva effettuata una necroscopia per esaminare accuratamente i vari organi. Una volta completata questa fase veniva raccolto un campione di parenchima renale, immerso in formalina e stoccato temporaneamente in frigorifero in attesa dell’esame istologico.
• CELIOSCOPIA
La celioscopia e la successiva biopsia renale sono state applicate con successo in tutti i soggetti che presentavano poliuria o oliguria. Il soggetto, sotto anestesia, era posizionato sul letto operatorio in decubito laterale destro, con le ali estese dorsalmente e doppiamente