Capitolo 2
Stress da sommersione
La sommersione è uno fra gli stress abiotici più dannosi per le specie vegetali in quanto impedisce l’assorbimento di ossigeno atmosferico attraverso le foglie e le radici a causa della sua bassa diffusione all’interno dell’acqua. La principale conseguenza è la scarsa respirazione da parte della pianta che porta ad una bassa produzione di energia e alla eventuale morte di cellule e tessuti che non sono in grado di adattarsi a tali condizioni di stress, in quanto il metabolismo ossidativo rappresenta il primo mezzo di rifornimento di energia per la crescita, lo sviluppo e le altre varie funzioni fisiologiche (Jackson e Armstrong, 1999).
Tutto ciò ha ovviamente un’enorme impatto soprattutto per le coltivazioni di interesse agronomico.
Generalmente le specie vegetali mostrano un diverso grado di tolleranza alla sommersione, tanto che possono essere classificate come idrofite, tolleranti o sensibili. Le risposte di queste diverse classi di piante comprendono cambiamenti temporanei dell’espressione genica e del metabolismo, ma anche cambiamenti anatomici e morfologici (Colmer et al., 1998; Richard et al., 1998; Jackson e Armstrong, 1999).
L’impatto negativo della sommersione sulla vita delle piante è una conseguenza della bassa velocità di diffusione dei gas nell’acqua rispetto all’aria e della bassa solubilità dell’O2 nell’acqua (Jackson, 1985; Armstrong e Drew, 2002). Nei tessuti non fotosintetici, come le radici, la [O ] diminuisce fortemente in seguito alla
sommersione, mentre si assiste ad un accumulo di CO2 prodotta durante la respirazione e la fermentazione (Armstrong et al., 1994; Crawford, 1992) . Nel caso invece di tessuti fotosintetici sommersi, questi sono caratterizzati da una variazione giornaliera significativa nella composizione dei gas (Raskin e Kende, 1984a):
- durante la fase luminosa la fotosintesi esaurisce la [CO2] endogena, mentre [O2] viene mantenuta costante al 21%;
- durante la fase buia la concentrazione di O2 endogeno è ridotto come risultato della respirazione, ma non vi è mai una totale riduzione perché l’acqua circostante ovviamente contiene O2 (Mommer et al., 2004). Un terzo gas, prodotto sempre endogenamente, è in grado di variare la propria concentrazione quando i tessuti della pianta sono sottoposti a periodi di sommersione. Si tratta dell’ormone vegetale etilene, il cui intrappolamento fisico associato ad un basso tasso di catabolismo nei suoi confronti (Hall, 1991) comporta un incremento della sua concentrazione all’interno dei tessuti sommersi. Un incremento fisiologicamente rilevante può avvenire addirittura entro pochi minuti dalla sommersione (Vreeburg et al., 2005).
Nonostante la capacità nel rispondere a rapidi e continui cambiamenti, alcune specie vegetali riescono a tollerare e sono in grado di crescere vigorosamente in condizioni di sommersione. Si tratta comunque di specie provenienti da ambienti paludosi, anche se non tutte sono in grado di tollerare periodi di completa sommersione (Blom e Voesenek, 1996).
Uno dei principali problemi incontrati dalle piante in tali condizioni è il deficit energetico causato dalla riduzione di O2 che appunto comporta l’inibizione dell’attività respiratoria cellulare. Dunque acclimatazioni di tipo metabolico consentiranno di continuare a produrre energia nonostante le condizioni ipossiche sfavorevoli. Come già precedentemente detto, Arabidopsis mostra una forte induzione dei geni codificanti per enzimi coinvolti nel metabolismo degli zuccheri, nella glicolisi e nella via fermentativa quando la concentrazione di O2 scende a valori tali da sottoporre i tessuti a condizioni ipossiche e anossiche (Klok et al., 2002; Loreti et al., 2005; Liu et al., 2005).
Altri geni indotti in condizioni di sommersione, quindi in generale da livelli ipossici, sono quelli codificanti per una emoglobina non simbiotica, coinvolta nell’omeostasi dell’ossido nitrico e che gioca un ruolo importante nella rigenerazione di NAD+ per mantenere attiva la glicolisi (Dordas et al., 2004).
Un altro gruppo di geni indotti è infine quello associato con la detossificazione delle ROS: perossidasi, ascorbato perossidasi, monoidroascorbato, glutatione riduttasi e superossido dismutasi (Klok et al., 2002). Molto probabilmente i prodotti di questi geni giocano un ruolo chiave nella protezione da stress ossidativi in post anossia.
Una seconda strategia messa in atto dalle piante in condizioni di sommersione è associata ad un adattamento a lungo termine e si tratta in genere di cambiamenti che riguardano l’anatomia e la morfologia fogliare, la formazione di nuovi tessuti (aerenchima) e l’allungamento di particolari organi come ad esempio i piccioli (Voesenek et al., 2006). Si tratta delle cosiddette strategie di “fuga” che consentono di ristabilire un contatto fra i tessuti sommersi e l’atmosfera sopra la superficie dell’acqua, in modo da consentire alla pianta di ristabilire l’attività metabolica aerobica (Voesenek et al., 2006; Kende et al., 1998). Un bilancio tra costi e benefici derivanti da una strategia di questo tipo suggerisce che le piante adottano la “fuga” come meccanismo di adattamento solo in caso di brevi periodi di sommersione (Perata e Voesenek, 2007).
Nel caso invece di sommersione per periodi prolungati o situazioni in cui questa avvenga a profondità tali da impedire che gli organi della pianta in allungamento entrino in contatto con l’atmosfera al di sopra della superficie acquosa, il rapporto tra costi energetici e benefici diventa sfavorevole per la pianta che invece adotterà una strategia “quiescente” caratterizzata da una crescita rallentata, che consente una maggiore conservazione di energia e carboidrati (Fukao e Bailey-Serres, 2004).
Per quanto riguarda il meccanismo molecolare alla base del modello “quiescente” di adattamento, due diversi gruppi di ricerca (Fukao et al., 2006; Xu et al., 2006) hanno ipotizzato che alla base delle variazioni in termini di tolleranza alla sommersione fra le diverse cultivar di riso, vi sia un putativo fattore di risposta per l’etilene (ethylene response factor, ERF).
2.1 Adattamenti morfologici e anatomici: le strategie di
“fuga” dalla sommersione.
Una delle specie vegetali maggiormente usata per studiare i meccanismi di acclimatazione e di “fuga” durante periodi di completa sommersione è Rumex
palustris. Si tratta di una pianta semi-acquatica appartenente al genere di Angiosperme dicotiledoni della famiglia delle Poligonacee, ampiamente diffusa nelle regioni temperate dell’emisfero boreale.
Figura 2.1: Movimento iponastico in R. palustris. (Voesenek et al., 2003)
• Anatomia e morfologia fogliare: le foglie di R. palustris hanno la parete cellulare
dell’epidermide e una cuticola estremamente sottili e i cloroplasti sono orientati verso l’epidermide anziché verso gli spazi intercellulari (Mommer et al., 2005b). La diffusione attraverso la cuticola potrebbe essere un metodo efficiente per la CO2 per penetrare all’interno delle foglie sommerse (Frost-Cristensen et al., 2003) anche nel caso in cui le foglie siano dotate di stomi. Sebbene non sia ancora del tutto chiaro il movimento degli stomi in piante sottoposte a sommersione, è noto che in Rumex palustris essi sono sicuramente chiusi quando la pianta è completamente sommersa (Mommer et al., 2005b). In risposta a periodi di sommersione prolungati alcune specie vegetali sono in grado di dare origine a nuove foglie acclimatate in acqua in grado di aumentare gli scambi gassosi con la superficie di acqua (Mommer e Visser, 2005). Ad esempio, in Rumex le foglie più giovani sono in grado di allungarsi oltre lo strato di acqua che le sovrasta e sebbene la maggior parte di tale allungamento avviene esclusivamente a livello del picciolo, anche la lamina fogliare è in grado di allungarsi e di diventare più stretta. In una fase successiva invece esse diventano più sottili e ciò consente un migliore scambio di gas tra la foglia e l’ambiente circostante (Mommer et al., 2005a). Questi cambiamenti morfologici e anatomici nelle giovani foglie acclimatate consentono un aumento dell’ingresso di CO2 in modo tale che possa essere mantenuta su livelli sufficientemente alti l’attività fotosintetica per garantire un approvvigionamento di zuccheri alla pianta.
• Formazione dell’aerenchima: l’aerenchima è un particolare tessuto che consente
un interconnessione di tipo longitudinale tra i vari organi della pianta in modo tale da permettere il trasporto dei gas lungo questa via. Si forma nelle radici a livello della corteccia mentre nel fusto avviene sia nella corteccia che nel midollo (Armstrong, 1979). Il volume occupato dall’aerenchima formato dipenderà essenzialmente dal genotipo in questione e dalle condizioni ambientali presenti (Colmer, 2003).
Sono stati individuati due diversi tipi di aerenchima in base al tipo di formazione (Sachs, 1882):
- “schizogenus” aerenchima, che si origina a partire dalla crescita differenziale delle cellule adiacenti la cui separazione avviene a livello della lamella; - “lisygenous” aerenchima, che invece è ottenuto dal collasso delle cellule grazie
all’azione dell’ormone etilene, il segnale principale che porta alla morte cellulare programmata (PCD) specialmente nelle cellule corticali che saranno così rimosse (Drew et al., 2000).
Nella maggior parte delle piante che provengono da zone paludose l’aerenchima si forma costitutivamente, ma è in particolar modo la risposta alla sommersione che induce la formazione di questo tessuto. Infatti nella maggior parte delle piante che non provengono da zone paludose l’aerenchima è quasi sempre assente e la sua formazione è indotta da particolari condizioni ambientali, come appunto la sommersione (Armstrong, 1979; Colmer, 2003), dalla compattazione del terreno o dalla carenza di nutrienti (Drew et al., 1989; He et al., 1996).
Non tutte le cellule però rispondono al segnale dell’etilene, per cui rimangono intatte e sono in grado di mantenere le connessioni simplastiche fra lo stele e il rizoderma. Il meccanismo per cui solo alcune cellule sono in grado di rispondere al segnale da etilene non è ancora del tutto conosciuto (Evans, 2003), anche se alcuni aspetti della via di traduzione del segnale, eventi che riguardano la modificazione dell’ultrastruttura cellulare che portano alla PCD e altri aspetti biochimici coinvolti sono stati descritti. Un modello proposto da Drew (2000) suggerisce che in questa via di segnale mediata dall’etilene vi siano a valle proteine, non ancora note, che vengono fosforilate in modo tale da indurre variazioni nell’espressione di quei geni che controllano il meccanismo PCD nelle cellule target. Ad esempio nel caso di formazione di aerenchima lisogeno in radici di Z. mays sono stati individuati sia
cambiamenti a livello ultrastrutturale, sia nell’espressione di alcuni enzimi, in particolare cellulasi, che hanno un ruolo chiave nella degradazione della parete cellulare durante la formazione di aerenchima (He et al., 1994; Saab e Sachs, 1996).
• Allungamento del picciolo: fra i cambiamenti anatomici, morfologici e di crescita
che avvengono quando la maggior parte delle specie vegetali subisce una parziale o totale sommersione, quelli che hanno suscitato un maggiore interesse sono stati l’alterata orientazione dei piccioli da una posizione orizzontale rispetto al terreno ad una verticale (definita crescita iponastica) e la forte induzione dell’allungamento dei piccioli e degli internodi. Si tratta di modificazioni che sono strettamente dipendenti dallo stadio di sviluppo della pianta (Groenveld e Voesenek, 2003).
Una delle prime modificazioni che avvengono in alcune specie vegetali in seguito a sommersione è l’incremento dell’angolatura dei piccioli più giovani rispetto al terreno, che consente di ridurre la distanza fra la foglia sommersa e la superficie acquosa (Banga et al., 1997). Si tratta di una tipica risposta indotta da sommersione che è stata individuata e studiata in diverse specie vegetali come Ranunculus repens, Caltha palustris, Leontodon taraxacoides, Paspalum dilatatum, R. palustris e A. thaliana (Voesenek et al., 2006). Altri fattori sono comunque in grado di determinare questo tipo di risposta, come ad esempio l’ombreggiamento e le alte temperature (Ballarè, 1999; Pierik et al., 2004; Millenar et al., 2005). Nel caso specifico di R. palustris questo tipo di risposta, definita crescita iponastica, è dovuta ad una differenziale crescita delle cellule a livello del picciolo, in quanto le cellule epidermiche che si trovano sulla superficie abassiale si allungano molto più velocemente rispetto a quelle che si trovano invece in posizione adassiale, determinando così la tipica curvatura verso l’alto delle foglie (Cox et al., 2004). Un prerequisito fondamentale affinché tale fenomeno si possa osservare è che l’angolatura minima dei piccioli al momento della sommersione sia di almeno 40° - 50° rispetto al terreno. Al di sotto di questa soglia minima risulta molto difficile osservare il fenomeno dell’allungamento (Cox et al., 2003).
Sicuramente il principale segnale in grado di far avvenire una serie di reazioni a cascata che portano all’allungamento delle cellule è l’accumulo di etilene. Un effetto diretto dell’aumento della concentrazione di etilene è la rapida
diminuzione del rapporto tra altri due ormoni vegetali, ovvero l’acido abscissico (ABA) e le gibberelline (GA), dovuto in particolar modo ad una netta diminuzione dei livelli di concentrazione di ABA endogeno (Benschop et al., 2005) contrapposta all’incremento di GA endogeno (Rjinders et al., 1997). Sia per Rumex che per riso è noto che ABA sia un regolatore negativo dell’allungamento cellulare, ma come ciò sia regolato a livello fisiologico non è ancora del tutto chiaro. Recenti studi hanno comunque dimostrato che ABA interferisce direttamente con la via biosintetica delle gibberelline, inibendo in questo modo l’attività promotrice della crescita da parte di GA (Benschop et al., 2005). Sempre da questi risultati è stato infine osservato che la riduzione dei livelli di ABA non associata ad incrementi della concentrazione di etilene non può essere sufficiente per determinare la risposta di allungamento del picciolo. Oltre alle variazioni delle concentrazioni ormonali, affinché vi sia un allungamento cellulare per poter ottenere una risposta iponastica è importante che vi sia la rottura delle pareti cellulari, assorbimento di acqua e sintesi di polisaccaridi per la parete cellulare (Cosgrove, 1999). Tutto ciò risulta associato, molto probabilmente, all’azione di particolari proteine di parete appartenenti alla famiglia delle espansine (McQueen-Mason et al., 1992). L’attività delle espansine presenta un proprio optimum di pH intorno a valori sub-acidi ed è stato ipotizzato che siano in grado di rompere i legami a idrogeno tra le emicellulose e le microfibrille di cellulosa. La loro espressione genica e l’attività proteica in molte specie vegetali sono strettamente correlate ai fenomeni di allungamento cellulare indotti da sommersione (Vreeburg et al., 2005).
Studi condotti sempre su Rumex dimostrano che i livelli di concentrazione di etilene sono alla base di due vie parallele di trasduzione del segnale: quella che porta alla variazione del rapporto ABA:GA e quella che invece porta all’induzione dei geni per le espansine e il conseguente aumento della concentrazione delle espansine a livello proteico (Vreeburg et al., 2005). Entrambe le vie consentono dunque il fenomeno dell’allungamento cellulare determinando così la tipica risposta iponastica in molte specie in conseguenza alla sommersione.
2.2
Adattamenti alla sommersione in specie vegetali di
particolare interesse agronomico: il ruolo di Sub1a in riso
come strategia “quiescente”.
Il riso rappresenta una fonte essenziale di cibo per miliardi di persone. Da un punto di vista agronomico la sua importanza è dovuta in particolar modo all’abilità di questa coltura di tollerare diverse condizioni ambientali avverse. Ad esempio il riso è una delle poche colture in grado di germinare e crescere in terreni costantemente sommersi (Alpi e Beevers, 1983; Perata e Alpi, 1993) per la sua capacità di rispondere a condizioni avverse di questo tipo attraverso determinati adattamenti morfologici e anatomici, come la formazione di aerenchima in grado di garantire la diffusione di O2 dagli organi aerei fino ai tessuti sommersi della pianta. Mentre la germinazione in anossia è ottenuta per mezzo di adattamenti di tipo metabolico, la formazione dell’aerenchima e altre modificazioni morfologiche non sono altro che strategie di fuga che consentono, per mezzo dell’ossigenazione, un rapido allungamento cellulare che permetta agli organi sommersi di raggiungere la superficie acquosa (Perata e Voesenek, 2007). Tuttavia in condizioni di completa sommersione la maggior parte delle cultivar di riso non sono in grado di tollerare questo tipo di stress, almeno non oltre una settimana (Xu et al., 2006).
Fra le poche cultivar in grado di tollerare invece lunghi periodi di completa sommersione (fino a due settimane), ricordiamo FR13A. La sua tolleranza alla sommersione è dovuta ad un locus genico, noto come Submergence1 (Sub1), localizzato sul cromosoma 9 (Xu e Mackill, 1996). Studi recenti (Xu et al., 2006) hanno dimostrato che la regione Sub1 di FR13A codifica per tre fattori di trascrizione (Sub1A, Sub1B e Sub1C) appartenenti al sottogruppo B-2 degli “ethylene response factor” (ERFs). La trascrizione di Sub1A e di Sub1C è fortemente regolata dalla sommersione, ma Sub1C è meno indotto in presenza di Sub1A, suggerendo quindi che Sub1A potrebbe reprimere Sub1C. Il terzo fattore di trascrizione (Sub1B) è invece lievemente regolato dalla sommersione. In termini di tolleranza alla sommersione è stato osservato che questa risulta essere fortemente correlata con la presenza dell’allele Sub1A-1 mentre la mancata tolleranza è associata o alla presenza dell’allele Sub1A-2 oppure all’assenza del gene Sub1A (Fukao et al., 2006). Questi dati sono confermati dai risultati ottenuti dalla trasformazione della varietà japonica, intollerante alla sommersione, con l’allele Sub1A-1 che appunto conferisce tolleranza alle piante transgeniche ottenute (Xu et al., 2006).
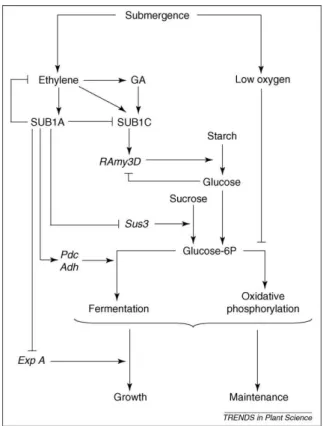
Sulla base di questi risultati è stato proposto un modello secondo cui l’incremento dei livelli di etilene determina un accumulo dei trascritti di Sub1A nelle cultivar di riso tolleranti la sommersione. Si tratta di una strategia quiescente che consente la sopravvivenza alla sommersione in quanto la proteina SUB1A incrementa la trascrizione dei geni associati alla fermentazione etanolica e reprime la trascrizione dei geni associati all’allungamento cellulare e al catabolismo dei carboidrati. Inoltre SUB1A è responsabile di una inibizione a feedback nella produzione di etilene.
Sub1 consente di aumentare la tolleranza alla sommersione se introdotto in cultivar di riso non tolleranti e in condizioni di crescita “normali” (non sommersione) non comporta nessun effetto negativo in termini di sviluppo, di indice di raccolta e qualità del seme (Xu et al., 2006).
Fig. 2.2: Modello riassuntivo che pone in relazione Sub1A con la tolleranza alla sommersione (Perata
e Voesenek, 2007).
2.3 La percezione del segnale di sommersione: cambiamenti
fisico-chimici del suolo.
Per capire come le piante rispondono o si adattano alla sommersione è importante conoscere il comportamento del sistema radicale e il suolo in cui esso si trova. Come
e quando lo stress sia percepito dalle radici non è ancora chiaro, ma è stato comunque ipotizzato che gli eventi spazio-temporali dei segnali potrebbero essere correlati con le alterazioni del suolo dovute alla sommersione (Dat et al., 2004). Sicuramente tutte le alterazioni che avvengono nella rizosfera potrebbero avere un ruolo nella percezione dello stress, ma individuare fra essi i potenziali segnali risulta difficile. Alternativamente individuare la sequenza temporale dei cambiamenti fisico-chimici del suolo e correlarli con le risposte delle piante potrebbe essere utile per conoscere gli ipotetici segnali (Dat et al., 2004).
Il primo evento segnale è dato dalla saturazione del suolo dovuto all’incremento dei livelli di acqua. Le cellule dell’apparato radicale non sembrano essere in grado di percepire direttamente i cambiamenti dei livelli di H2O, ma l’identificazione di una istidina-kinasi transmembrana in Arabidopsis (Urao et al., 1999) potrebbe far supporre che essa possa svolgere il ruolo di osmosensore dei livelli di H2O. Risulta ormai noto che le cellule siano in grado di utilizzare diversi “indicatori” per la disponibilità di H2O (variazioni nella pressione di turgore, nel volume cellulare o area di membrana oppure cambiamenti nelle connessioni parete-membrana) come possibili elementi di percezione, ma poiché la saturazione del suolo altera decisamente il potenziale osmotico e la pressione di turgore dopo appena due ore (Jackson et al., 1996), è ipotizzabile che questo osmosensore abbia un ruolo chiave nella percezione e trasmissione del segnale (Dat et al., 2004).
Un altro cambiamento fisico-chimico che può rappresentare un segnale di stress da sommersione è la bassa disponibilità di ossigeno, in quanto è noto che questo diffonde in acqua molto più lentamente che in aria. Ciò ovviamente comporterà una riduzione nella produzione di ATP con conseguente riduzione delle attività metaboliche a livello radicale. Sicuramente la bassa disponibilità energetica può di conseguenza ridurre altri processi cellulari come l’uptake di sostanze nutrienti, la regolazione del pH citoplasmatico o aggiustamenti di tipo osmotico (Dat et al., 2004).
Altre variazioni a livello chimico in seguito a sommersione riguardano il pH del suolo e il potenziale redox. Tali modificazioni potrebbero avere un effetto non diretto sulla crescita della pianta, ma ad esempio potrebbero ridurre la disponibilità di determinati elementi minerali comportando un riduzione dell’attività fotosintetica con conseguente alterazione del metabolismo e della crescita della pianta (Dat et al., 2004).
La bassa diffusione dei gas in H2O potrebbe portare all’accumulo di co-prodotti del metabolismo anaerobico potenzialmente fitotossici come etanolo, acido lattico, CO2, N2, H+ e metano. Questi potrebbero accumularsi a livello intracellulare o essere rilasciati nel suolo alterandone così le proprietà chimiche, diventando così ipotetici segnali dello stress.
2.4 Attività fotosintetica in condizioni di sommersione
L’attività fotosintetica in condizioni di sommersione è piuttosto limitata a causa della bassa disponibilità di luce e di CO2 (Sand-Jensen, 1989). La luce che raggiunge le foglie delle piante sommerse è attenuata dall’acqua e dalla materia organica presente, da particelle tipo limo o dalle alghe presenti sulla superficie acquosa (Vervuren et al., 2003).
Le concentrazioni di CO2 negli organi sommersi risultano essere molto più basse rispetto a quelle normalmente richieste per mantenere alta l’attività fotosintetica (Sand-Jensen, 1989). Durante la sommersione avvengono comunemente fluttuazioni giornaliere dei livelli di CO2 con un accumulo durante la fase buia ed un rapido abbassamento quando l’intensità luminosa è in grado di far avviare l’attività fotosintetica. La capacità di assorbimento della CO2 da parte delle foglie sommerse limita sicuramente la fotosintesi a causa della lenta diffusione dei gas nell’acqua, per cui questo gas sarà un fattore limitante anche per le foglie acclimatate delle piante acquatiche (Raven et al., 1985). Inoltre per le piante terrestri la cuticola rappresenta un vero e proprio ostacolo per l’assorbimento di CO2 impedendo così l’attività fotosintetica nelle piante sommerse (Mommer et al., 2005b) determinando quindi un incremento della fotorespirazione (Mommer et al., 2005b).
La composizione e la variazione dei livelli dei gas prodotti endogenamente, in particolare di O2, CO2, ma anche dello stesso etilene, rappresentano indubbiamente delle potenziali molecole segnale dello stress da sommersione. Anche la disponibilità della luce ricopre un importante ruolo dal momento che, come è stato appena detto, influenza le concentrazioni interne di O2 e CO2 agendo sull’attività fotosintetica e sulla respirazione.
Dal momento che questi due gas sono direttamente influenzati dalla qualità e intensità luminosa, l’etilene viene considerato come segnale più attendibile dello stress da sommersione (Voesenek e Blom, 1999).
Oltre a questi tre gas, anche le ROS (reactive oxygen species), che si accumulano proprio durante periodi di sommersione, svolgono un ruolo nei meccanismi di regolazione coinvolti nell’adattamento a questo stress (Bailey-Serres e Chang, 2005).