Indice
Riassunto
pag. 2
Introduzione
pag. 4
La corteccia parietale posteriore (PPC)
pag. 7
La corteccia cingolata anteriore (ACC)pag. 11
Il cervelletto
pag. 15
Scopo dello studio
pag. 23
Materiali e metodi
pag. 25
Risultati
pag. 32
Discussione
pag. 45
Bibliografia
pag. 51
Riassunto
I territori corticali della regione frontale mediale fanno parte di un sistema di monitoraggio dell’azione in grado di rilevare e elaborare i segnali di errore.
Un correlato elettroencefalografico della attivazione di queste aree è il ritmo θ (theta). L’attività θ frontale mediale aumenta durante l’esecuzione di compiti complessi, quando è richiesto il monitoraggio continuo dell’azione e dopo la comparsa di una risposta errata.
È stato proposto che l’attivazione del sistema delle cortecce frontali mediali segnali la comparsa di un conflitto fra il risultato desiderato di una azione e il risultato ottenuto, valutabile attraverso il feedback sensoriale o prevedibile ancor prima che sia disponibile il feedback, sulla base della copia efferente del comando motorio.
Per verificare l’ ipotesi che la corteccia frontale mediale sia implicata nel rilevamento ed elaborazione dell’errore in compiti di integrazione visuomotoria si sono studiate, in un gruppo di soggetti normali, le variazioni di potenza spettrale e di coerenza nella banda di frequenza θ della corteccia frontale mediale durante l’esecuzione di un compito (task) di tracking1. Tale compito richiedeva al soggetto il confronto continuo tra il risultato atteso dell’azione e il risultato ottenuto e permetteva di correlare le variazioni dell’attività θ con l’errore commesso. Per paragone i soggetti eseguivano anche altri tre task che non richiedevano integrazione visuomotoria: un task puramente motorio (M), un task puramente visivo (V) e un task motorio associato a stimoli visivi distraenti (V+M). I segnali elettroencefalografici (EEG) registrati durante i diversi task sono stati sottoposti ad analisi delle componenti indipendenti, una tecnica capace di separare le diverse sorgenti di segnale EEG comunemente sovrapposte a livello dei singoli elettrodi e di localizzarle in aree corticali di interesse. In particolare, la nostra attenzione si è focalizzata su due sorgenti: una localizzata a livello della corteccia frontale mediale (IC1), l’altra a livello corteccia parietale posteriore mediale (IC6). Rispetto alla condizione di riposo, la potenza spettrale nella banda θ della corteccia frontale mediale (fmθ) aumentava significativamente durante il task VM indipendentemente dalla mano (dominante o non dominante) usata per eseguire il task. L’analisi comparata dei risultati ottenuti durante VM con quelli ottenuti negli altri tipi di task (M, V, V+M), in cui non si osservava alcuna significativa modifcazione del ritmo
1 Compito motorio nel quale il soggetto deve inseguire per mezzo di un cursore il percorso tracciato da un
θ, ha permesso di ascrivere gli aumenti di fmθ, osservati durante VM, al processo di integrazione visumotoria. Un risultato di particolare interesse è che l’aumento di fmθ durante VM si associava positivamente con l’errore commesso e, nel caso di esecuzioni particolarmente degradate, anche con un aumento di coerenza nella banda θ fra la corteccia frontale mediale e la corteccia parietale mediale.
I risultati sopra descritti sono in linea con l’ipotesi formulata originariamente che fmθ sia associato al monitoraggio continuo del movimento e più precisamente all’elaborazione e/o correzione dell’errore. Poiché anche il cervelletto partecipa alla generazione di un segnale di errore interno in caso di conflitto fra predizione ed esecuzione dell’azione, è stata intrapresa una seconda serie di esperimenti in pazienti cerebellari che eseguivano lo stesso tipo di task per verificare l’ipotesi che il cervelletto influenzi l’attività di elaborazione e/o correzione dell’errore a livello della corteccia frontale mediale.
Al contrario dei soggetti normali, i pazienti cerebellari non presentavano alcun aumento significativo di attività θ frontale mediale durante l’esecuzione del task VM, indipendentemente dalla mano usata e non mostravano alcuna associazione tra fmθ e indice di errore. Inoltre, l’analisi dello spettro di coerenza non ha mostrato alcun aumento significativo di coerenza tra la corteccia frontale mediale (IC1) e la corteccia parietale mediale (IC6) durante il task VM nè ha evidenziato alcuna modulazione della coerenza da parte dell’indice di errore.
In conclusione i risultati dello studio confermano ed estendono i dati della letteratura nel dimostrare che:
1) l’attività θ frontale mediale, putativamente originata nella corteccia cingolata anteriore, è associata al monitoraggio continuo dell’azione e più precisamente al rilevamento ed elaborazione on-line dell’errore;
2) questa associazione è quasi totalmente assente nel soggetto cerebellare.
Sulla base di questi risultati si può ipotizzare che in caso di conflitto tra predizione ed esecuzione il cervelletto influenzi il corretto rilevamento ed elaborazione dell’errore a livello della corteccia frontale mediale.
Introduzione
Fin dalla fine del 1800 i meccanismi che intervengono nel processo di integrazione sensori-motoria sono stati oggetto di estese ricerche. Dopo più di un secolo di studio molti aspetti sono stati chiariti, ma altri sono tuttora oggetto di controversia.
In particolare, è stato chiarito che il movimento dell’arto sotto controllo visivo, che è un caso particolare di integrazione sensori-motoria, è sotto il controllo di un mosaico di aree corticali reciprocamente interconnesse distribuite nei lobi frontale e parietale. Anche il concetto, elaborato più di una decina di anni fa da Goodale e Milner (1992), della distinzione anatomo-funzionale di due proiezioni a partenza dalle aree corticali visive primarie, una ventrale, che proietta alla corteccia infero-temporale, deputata alla percezione (“vision-for-perception”) ed una dorsale, che proietta alla corteccia parietale posteriore, deputata al controllo visuomotorio (“vision-for-action”), è andato incontro ad una rielaborazione. Un’interpretazione alternativa è che anzichè di distinzione fra percezione ed azione si debba parlare di un’organizzazione complessa con connessioni multiple visuomotorie che funzionano in parallelo (Pisella et al., 2006). Di queste connessioni almeno due associano aree parietali ed aree frontali: una proiezione dorso-dorsale che coinvolge le porzioni più dorsali del lobo parietale e le aree premotorie del lobo frontale ed una proiezione ventro-dorsale che unisce le porzioni più ventrali del lobo parietale alle aree prefrontali. La prima proiezione sarebbe implicata nel controllo visuomotorio on-line, mentre la seconda interverrebbe prevalentemente nella pianificazione e programmazione del movimento.
Appare pertanto che lo studio dei meccanismi di integrazione visuomotoria non possa prescindere dall’integrazione di due concetti apparentemente contradditori, che il controllo visuomotorio è organizzato in maniera complessa e distribuita nel circuito fronto-parietale e che, tuttavia, le aree parietali e frontali e le relative connessioni presentano, almeno in parte, segregazione anatomica e specificità funzionale. E’ stato suggerito che la definizione degli obiettivi e la pianificazione del movimento richiedano la partecipazione cosciente del soggetto e comportino il coinvolgimento delle aree frontali. La costruzione dettagliata del movimento, invece, dipenderebbe da un sistema che opera in maniera implicita, che ha localizzazione prevalentemente posteriore e che genera il comando motorio appropriato sulla base del contesto e degli obiettivi forniti dal sistema anteriore.
Un concetto che è stato recentemente elaborato considera che atti comportamentali quali la percezione e l’azione derivino dalla sincronizzazione dell’attività di neuroni localizzati in una stessa area cerebrale (sincronizzazione intra-area o short-range) o in aree cerebrali diverse (sincronizzazioni inter-area o long-range; Bressler et al., 2001; Kelso et al., 1997; Tononi et al., 1998; Varela et al., 2001). Nel caso di un movimento richiedente l’integrazione sensori-motoria sincronizzazioni inter-area assocerebbero i neuroni deputati alla definizione degli obiettivi del movimento, alla pianificazione dell’azione e al monitoraggio del risultato (neuroni delle aree frontali) con quelli che presiedono all’elaborazione degli schemi sensori-motori (neuroni delle parietali e motorie), consentendo di tradurre in azione l’intenzionalità motoria mediante la scelta dello schema sensori-motorio più appropriato. Tali schemi sensori-motori sono rappresentazioni neurali che simulano la relazione esistente fra i comandi motori e le loro conseguenze sensoriali, permettendo di predire il comando motorio necessario per ottenere uno stato desiderato (“modello inverso”) oppure di predire le conseguenze sensoriali di un comando motorio (“modello diretto o a forward”).
La capacità di alcune strutture cerebrali di predire le conseguenze di un comando motorio è stata oggetto di estese ricerche che nelle pagine che seguono saranno ampiamente discusse per la relazione esistente fra predizione e errore motorio.
La predizione, infatti, consentirebbe di generare un segnale di errore allorché è confrontata col risultato desiderato. E’ stato ipotizzato, in particolare, che se il sistema rileva una coincidenza sufficiente fra il risultato desiderato e quello previsto, come si verifica generalmente nel caso di un atto motorio appreso, lo stato di risonanza dei neuroni corticali rimanga ristretto all’interno delle aree di elaborazione sensori-motoria implicita (cortecce motorie e parietali). Se, al contrario, il sistema rileva una discrepanza fra il segnale desiderato e quello previsto, tale da superare la soglia per la correzione automatica del movimento (come nel caso di movimenti non appresi), sarebbe facilitata la diffusione dei patterns di attività oscillatoria coerente in un territorio corticale più ampio includente le aree esecutive frontali (Andre e Arrighi, 2003). L’attività delle aree frontali, in questo caso, provvederebbe ad inibire gli schemi motori rivelatisi inappropriati e contribuirebbe a selezionare lo schema sensori-motorio più opportuno.
Un concetto altrettanto importante rispetto a quello di cooperatività parieto-frontale nel processo di integrazione sensori-motoria è quello di cooperatività fra strutture sotto-corticali e sotto-corticali. Nell’ambito delle strutture sotto-sotto-corticali un ruolo chiave è
esercitato dal cervelletto che è storicamente implicato nell’elaborazione dell’errore motorio ed è provvisto di capacità predittive. Inoltre, è stato recentemente rielaborato il concetto di circuito cerebello-corticale, assumendo che esso sia organizzato secondo canali (“loop”) paralleli che associano aree specifiche della corteccia cerebellare con aree specifiche delle corteccia cerebrale comprendenti non solo le aree motorie, come storicamente considerato, ma anche le aree frontali e parietali (Leiner H. e Leiner A., 1997; Schmahmann e Pandya, 1997; Middleton e Strick, 1998b).
Alla luce dei dati sopra riportati la trattazione seguente prenderà in considerazione in maniera analitica i seguenti punti:
- la generazione del segnale di errore nel corso di un compito di integrazione visuomotoria a livello parietale, con particolare riferimento alla corteccia parietale posteriore,
- il monitoraggio e/o correzione del movimento a livello frontale, con particolare riferimento alla corteccia cingolata anteriore e al ritmo theta frontale mediale,
- il ruolo del cervelletto nel controllo del movimento e nella generazione di un segnale di errore in cooperazione con le aree corticali parietali e frontali,
al fine di fornire un substrato anatomo-fisiologico all’ipotesi che il cervelletto influenzi l’attività di rilevamento e/o elaborazione dell’errore visuomotorio a livello corticale e,in particolare, a livello della corteccia frontale mediale.
La corteccia parietale posteriore (PPC)
La corteccia parietale posteriore (PPC) è una porzione di corteccia parietale suddivisa dal solco intraparietale in lobulo parietale superiore (SPL) e lobulo parietale inferiore (IPL) .
La tradizionale suddivisione funzionale della PPC in SPL, deputato prevalentemente a funzioni somato-sensoriali, e in IPL, implicato maggiormente nella visione, oggi non é più sostenibile. Entrambi i lobuli ricevono e combinano informazioni molteplici, tra cui quelle somato-sensoriali e visive, e le usano per la composizione dei comandi motori (Andersen e Buneo, 2002; Battaglia-Mayer et al., 2003).
Ai fini della tesi la trattazione seguente sarà limitata ad alcune evidenze che dimostrano che la PPC ed in particolare SPL è sede di schemi sensori-motori e che questi schemi possono essere usati per il controllo implicito dell’azione e per la correzione del movimento. Come accennato precedentemente, si intende per schema sensori-motorio una rete neurale capace di produrre il comando motorio appropriato sulla base del risultato desiderato, o viceversa capace di predire le conseguenze sensoriali dell’atto motorio sulla base del comando dato.
Le modalità di risposta dei neuroni della PPC sono tali da rendere questa corteccia particolarmente adatta a contenere gli schemi sensori-motori. Questi neuroni, infatti, rispondono ad una molteplicità di segnali sia di natura sensoriale (visivi, uditivi, e vestibolari) che di natura motoria ed esibiscono inoltre selettività spaziale.
Per valutare la capacità della PPC di elaborare gli schemi sensori-motori e di usarli in maniera implicita per il controllo on-line del movimento Pisella et al. (2000) hanno valutato la capacità di soggetti normali di inibire la risposta correttiva ad un improvviso cambiamento di localizzazione del bersaglio. Il razionale dello studio risiedeva nell’osservazione che una volta che un movimento ballistico di raggiungimento di un bersaglio è stato programmato esso può essere corretto senza necessità di programmare un nuovo movimento qualora, ad esempio, il bersaglio sia improvvisamente spostato. In aggiunta, questa correzione si verificava indipendentemente dal fatto che il soggetto fosse consapevole dello spostamento del bersaglio. Dimostrando che i soggetti spesso non riuscivano ad evitare la correzione della traiettoria, gli Autori concludevano che la PPC dovesse contenere un “pilota automatico”, basato sulle capacità visuospaziali dei suoi neuroni, che presiedeva ad un meccanismo di controllo on-line indipendente dal controllo motorio intenzionale. Per confermare questa ipotesi, nella seconda parte dello studio di Pisella et al., lo stesso protocollo era applicato ad un paziente con lesione
bilaterale della PPC e con un quadro clinico di atassia ottica bilaterale. Sebbene il paziente fosse in grado di raggiungere un bersaglio fisso con relativa accuratezza, esso presentava un pattern anomalo di correzione della traiettoria quando la localizzazione del bersaglio cambiava improvvisamente: la correzione automatica, osservata nei soggetti normali, era soppressa, mentre i movimenti di correzione lenti ed intenzionali erano preservati. Il risultato di questo studio è in accordo con quello ottenuto da Desmurget et al. (1999), utilizzando la TMS (stimolazione transcranica magnetica) su soggetti normali per ottenere l’inattivazione funzionale della PPC ed un protocollo simile che richiedeva al soggetto di raggiungere un bersaglio che si spostava nel corso del movimento. In accordo col risultato dello studio precedente, l’applicazione della TMS sull’area distribuita intorno al solco intraparietale all’inizio del movimento impediva al soggetto di correggere la direzione del movimento.
Le conclusioni di questi studi sono che la PPC è necessaria per il controllo motorio on-line e per la correzione automatica del movimento, ma non per la sua programmazione, nè per il controllo motorio intenzionale.
La correzione automatica del movimento presuppone il rilevamento dell’errore. Evidenza che la PPC sia implicata nel rilevamento dell’errore è stata fornita da un recente esperimento (Tunik et al., 2005) in cui gli Autori hanno valutato se esistesse una finestra temporale durante la quale l’applicazione parietale della TMS fosse in grado di inibire la correzione del movimento. Se la stimolazione veniva applicata entro 65 msec dalla perturbazione, cioè prima dell’inizio della risposta correttiva essa inibiva la correzione; viceversa se la stimolazione veniva applicata dopo 65 msec questa risultava inefficace e la correzione procedeva indisturbata. Gli Autori concludevano quindi che la PPC ha un ruolo cruciale nella fase iniziale, quando viene rilevato l’errore, ma non è responsabile della fase di correzione.
L’ ipotesi che la PPC sia coinvolta nella elaborazione dell’errore è confortata anche dall’osservazione che essa presenta almeno tre proprietà che sono richieste a un sistema di rilevamento dell’errore. Innanzitutto, essa possiede le informazioni per localizzare sia il bersaglio che l’arto, ricevendo informazioni afferenti da vari canali sensoriali (visivo, propriocettivo, vestibolare) e dalle aree motorie principali (Andersen et al., 1997; Brodal e Bjaalie, 1997). Secondariamente, ha la capacità di riferire informazioni eterogenee (per es. i segnali relativi all’arto e al bersaglio) ad uno stesso sistema di riferimento (Clower et al., 1996; Carey et al., 1997; Binkofski et al., 1999; Xing e Andersen, 2000). Infine, è stato dimostrato che durante un movimento di
raggiungimento del bersaglio l’attività di alcuni gruppi di neuroni della PPC varia man mano che la mano si approssima al bersaglio (MacKay, 1992). L’attività di questi neuroni è quindi modulata dall’errore motorio. Pertanto, è stato suggerito che la PPC partecipi all’elaborazione dell’errore motorio provvedendo al confronto continuo e reciproco fra la localizzazione del bersaglio e quella della mano servendosi delle informazioni sensoriali periferiche (Desmurget et al., 1999; Della Maggiore et al., 2004).
Un ipotesi aggiuntiva ma non incompatibilie con la precedente è che il rilevamento dell’errore da parte della PPC sia basato sulle capacità predittive di questa area. Questa ipotesi presuppone che la PPC sia sede di un modello interno a forward. Come accennato precedentemente il modello interno è una rappresentazione interna del movimento capace di simulare il movimento naturale. Si distinguono modelli inversi ed a forward (Fig.II A e B). Dato lo stato attuale e lo stato desiderato di un sistema il modello inverso consente di elaborare il comando motorio necessario per ottenere lo stato desiderato. Un modello a forward al contrario, consente di stimare lo stato che un sistema assumerà dato lo stato corrente e il comando motorio applicato. E’ pertanto evidente che il modello a forward assolve a una funzione predittiva, permettendo al sistema nervoso centrale di predire il risultato di un comando motorio senza necessità di informazioni circa il movimento dalla periferia.
Come sopra accennato è stato recentemente suggerito che la PPC provveda al rilevamento dinamico dell’errore attraverso un meccanismo a forward (Desmurget e Grafton, 2000). In accordo con questa ipotesi, durante un movimento, ad es. del braccio verso un bersaglio, il modello a forward della dinamica dell’arto, consentirebbe la predizione della posizione finale dell’arto stesso. Se la localizzazione finale predetta dell’arto e la posizione del target non coincidono, sarebbe generato precocemente un segnale di errore.
Un certo numero di studi ha verificato sperimentalmente le capacità predittive della PPC (Dulhamel et al., 1992; Colby et al., 1996; Eskandar e Assad, 1999). A titolo di esempio, alcuni neuroni parietali visivi iniziano a scaricare in relazione ad uno stimolo luminoso prima ancora che si verifichi il movimento saccadico oculare che porterà lo stimolo luminoso nel loro campo recettivo. Ciò sta ad indicare che la corteccia parietale anticipa le conseguenze retiniche dei movimenti oculari (Dulhamel et al., 1992).
La capacità predittiva della PPC è stata confermata anche da studi neuropsicologici di rappresentazione mentale dei movimenti della mano in pazienti affetti da lesione di
questa area (Sirigu et al., 1996). Soggetti con lesione della PPC, al contrario dei soggetti normali o di pazienti con lesione della corteccia motoria primaria, non riuscivano a predire, mediante la rappresentazione mentale, il tempo necessario per eseguire movimenti di raggiungimento di un bersaglio o movimenti differenziati delle dita. Questi dati suggeriscono che la capacità di immaginare un movimento coinvolge un modello a forward che si basa sul comando motorio programmato (ma non eseguito) e che questo modello a forward dipende criticamente dall’integrità della PPC.
La corteccia cingolata anteriore (ACC)
La corteccia cingolata anteriore è una regione frontale posta al centro di tre importanti sistemi anatomo-funzionali: motorio, limbico e prefrontale (Paus, 2001; Wang et al., 2005). Le sue dense proiezioni alla corteccia motoria e al midollo spinale testimoniano il suo ruolo nel controllo motorio; le connessioni con l’ippocampo e la corteccia orbitale posteriore implicano il suo coinvolgimento nel sistema limbico; le afferenze dai nuclei mediali del talamo e dai nuclei del tronco encefalico consentono di associarla al controllo vegetativo; infine, le connessioni cortico-corticali reciproche con la corteccia prefrontale laterale forniscono il substrato anatomo-funzionale al noto ruolo della ACC nel controllo cognitivo. In accordo con queste evidenze anatomiche studi di neuroimmagine hanno dimostrato l’attivazione della ACC in una varietà di compiti che richiedono attenzione selettiva, elaborazione delle informazioni a contenuto emozionale e motivazionale, memoria di lavoro e selezione di risposte potenzialmente conflittuali. In un compito che valutava separatamente l’impegno attentivo rispetto all’apprendimento, condizioni entrambe in grado di attivare la ACC e la corteccia prefrontale dorsale, il grado di attivazione di queste due aree era maggiore quando era richiesto di apprendere un nuovo compito piuttosto che di focalizzare l’attenzione sul compito già appreso (Jueptner et al., 1997). Gli Autori concludevano che l’attivazione delle due aree frontali era prevalentemente, ma non esclusivamente, associata al monitoraggio del risultato del nuovo compito e alla necessità di selezionare e memorizzare delle sequenze motorie corrette.
L’ipotesi attualmente più accreditata è che la ACC sia attivata in compiti che richiedono al soggetto di scegliere fra risposte potenzialmente conflittuali e che monitori, pertanto, la competizione tra queste risposte (Carter et al., 1998; Botvinick et al., 2001; Van Veen et al., 2001). Questa ipotesi consente di estendere il noto concetto di associazione fra ACC ed errore delineando una funzione prevalentemente ”valutativa” di questa area nel controllo on-line dell’azione (Carter et al., 2000; Botvinick et al., 2004). Ai fini dell’attivazione della ACC l’errore diventa un particolare caso di conflitto che si stabilisce fra la risposta errata, che il soggetto deve inibire, e l’alternativa corretta, che deve rendere esecutiva (Van Veen e Carter, 2002).
Per ciò che concerne l’errore che compare nel corso di un compito di integrazione visuomotoria, studi di risonanza magnetica funzionale (fMRI) hanno dimostrato che la ACC è attivata nel corso di compiti visuomotori. Più precisamente, in uno studio fMRI di simulazione di guida in cui l’entità del segnale era correlata con la velocità di guida,
l’attivazione della ACC diminuiva con la riduzione della velocità (proporzionalmente cioè alla riduzione della richiesta di controllo), suggerendo che l’attivazione di questa area correli col monitoraggio on-line dell’azione (Calhoun et al., 2002).
In un altro studio sempre di simulazione di guida il segnale RM registrato in ACC correlava negativamente con la qualità dell’esecuzione del compito (Uchyama et al., 2003). In particolare in questo studio il soggetto doveva mantenere una certa distanza dalla macchina che lo precedeva e quindi rilevare e/o elaborare on-line l’errore (distanza tra le macchine) e doveva scegliere tra due risposte conflittuali (accelerazione /decelerazione).
Recentemente è stato dimostrato che durante l’apprendimento di un nuovo compito (associazione arbitraria di stimoli visivi e di risposte motorie) la responsività di un’area all’interno della ACC al feedback sensoriale o a un segnale di errore associato alla risposta, variava in funzione del grado di apprendimento (Mars et al., 2005). In accordo col concetto che in un compito totalmente nuovo l’informazione dell’errore dipende dal feedback sensoriale periferico piuttosto che da un sistema predittivo interno, l’attivazione della ACC associata al feedback errato era massima all’inizio e diminuiva nel corso dell’apprendimento. Al contrario, l’attivazione associata alla risposta errata, indice della generazione di un segnale di errore interno, era minima inizialmente ed aumentava nel corso dell’apprendimento. In conclusione, questi dati confermano che la ACC può rispondere a segnali di errore sia interni che esterni, come suggerito da numerosi studi (Carter et al., 1998, Ullsperger e Von Cramon, 2001 e 2003; Holroyd et al., 2004).
Un correlato elettroencefalografico (EEG) dell’attivazione della ACC è il ritmo θ frontale mediale (fmθ). Un numero consistente di studi ha localizzato l’attività θ registrata dalla corteccia frontale mediale nell’ambito della ACC o direttamente, associando la registrazione EEG a tecniche di neuroimmagine (Pizzagalli et al., 2003), o indirettamente, associando indipendentemente fmθ e attivazione della ACC a compiti di attenzione focalizzata, memoria di lavoro, elaborazione affettiva e risposte di orientamento (Davis et al., 1997; Gevins et al., 1997; Asada et al., 1999; Tesche et al., 2000; Aftanas et al., 2001).
Infine, è stato dimostrato che il ritmo θ registrato con elettrodi di superficie nella regione fronto-mediale corrisponde a una o più sorgenti di segnale (dipoli) localizzati nella porzione dorsale della ACC (Dehaene et al., 1994; Miltner et al., 1998; Ullsperger e von Cramon, 2001).
Sorprendentemente, data l’origine di fmθ nella ACC e l’associazione della ACC con l’errore/conflitto, un numero relativamente limitato di studi ha preso in considerazione il rapporto fra questo ritmo ed il monitoraggio on-line dell’azione.
Negli anni ’80-90 tre studi hanno riportato la comparsa di fmθ durante un compito visuomotorio e lo hanno associato al grado di difficoltà del compito (Mizuki et al., 1982) o alle caratteristiche dell’esecuzione (Lang et al., 1987; Laukka et al., 1995), riferendolo prevalentemente a concentrazione ed attenzione focalizzata.
Recentemente, due gruppi di Autori hanno parallelamente valutato l’associazione fra fmθ ed errore, localizzando la sorgente di segnale EEG a livello fronto-centrale mediante l’impiego dell’analisi delle componenti indipendenti. Entrambi i gruppi hanno dimostrato l’associazione fra fmθ ed errore o direttamente, registrando “burst” di attività θ durante o immediatamente dopo l’azione errata (Makeig et al., 2002), o indirettamente (Luu et al., 2004), dimostrando che l’attività θ rende conto, per oltre il 50%, dell’ampiezza del potenziale negativo che compare precocemente dopo una risposta errata in sede fronto-mediale (potenziale errore-correlato o ERN).
Fin dagli anni ‘90 a cui risalgono le prime descrizioni (Falkenstein et al., 1990 e 1991; Gehrin et al., 1993), l’ERN ha rappresentato il correlato elettrofisiologico cerebrale dell’errore. Da quando il corrispondente generatore è stato tentativamente localizzato nella porzione dorsale della ACC (e in parte nella corteccia prefrontale laterale) l’ERN, come la ACC, è stato associato al rilevamento (Falkenstein et al., 1990; Gehrin et al., 1993) e/o correzione dell’errore (Rodrìguez-Fornells et al., 2002), poi al conflitto fra due o più risposte contrastanti fino alla attribuzione di un ruolo più generale di indicatore del processo di monitoraggio on-line dell’azione. Per l’interpretazione del significato funzionale dell’ERN (e indirettamente dell’ACC) un dato di estrema importanza deriva dallo studio del suo sviluppo temporale. L’evidenza che il picco di questo potenziale compare troppo precocemente rispetto alla risposta (60-80 ms) per dipendere da informazioni sensoriali suggerisce che l’ERN rifletta un meccanismo di controllo basato sulla predizione (Bernstein et al., 1995). Questo meccanismo confronterebbe il risultato desiderato di un’azione con il risultato previsto, stimato sulla base del segnale fornito da un modello interno a forward a sua volta attivato dalla copia efferente del comando motorio. E’ stato proposto che ogni qualvolta il sistema rilevi un “mismatch” fra il risultato desiderato e quello previsto venga generato un segnale di errore interno capace di attivare la ACC la quale provvederebbe ad arrestare la risposta errata precocemente e a selezionare la risposta corretta il prima possibile. Appare quindi
che la ACC faccia parte di un sistema cerebrale a feedforward di monitoraggio on-line dell’azione.
A fronte di un elevato numero di studi neuro- o psicofisiologici che hanno valutato la relazione fra ERN ed errore in compiti prettamente coinvolgenti le funzioni esecutive, un solo studio elettrofisiologico, a nostra conoscenza, ha valutato l’associazione di ERN ed errore durante l’esecuzione di un compito di integrazione visuomotoria (Contreras-Vidal e Kerick, 2004). Questo compito induceva precocemente una negatività a livello di un cluster di componenti indipendenti con proiezione topografica medio-frontale, simile per sede di origine, sviluppo temporale e relazione con l’errore all’ ERN classicamente descritto. In particolare, il cluster era consistentemente presente sia quando i soggetti eseguivano il movimento (tracciare una linea retta dal centro alla periferia in varie direzioni) in presenza di normale feedback del movimento della penna (visualizzata come cursore su uno schermo) sia quando lo eseguivano in condizioni di feedback alterato (rotazione della posizione del cursore sullo schermo). E’ intuitivo che nel primo caso l’errore visuomotorio, dopo opportuno allenamento, era basso, mentre nel secondo caso era elevato, almeno finchè il soggetto non raggiungeva un opportuno adattamento. In aggiunta, l’esame dell’attività del cluster in funzione della frequenza mostrava un transitorio aumento dell’attività nella banda θ nel range temporale 200-500 ms rispetto all’inizio del movimento.
Dato che, oltre al cluster fronto-centrale erano identificati altri cluster a proiezione centro-posteriore e a transitoria attività θ in associazione temporale fissa con l’inizio del movimento, gli Autori concludevano che il compito visuomotorio è associato all’attivazione di una rete neurale a distribuzione fronto-parietale, e formulavano l’ipotesi secondo cui l’area fronto-centrale provvede al monitoraggio dell’azione e a generare “un segnale” per il reclutamento di strutture fronto-parietali addizionali in caso di errore visuomotorio.
Il cervelletto
Il cervelletto è una struttura sottocorticale con funzioni complesse concernenti
classicamente il processo di integrazione sensori-motoria ed il controllo del movimento con particolare riferimento ai parametri spaziali e temporali (Stein, 1986; Horne e Butler, 1995). L’esecuzione di un compito di tracking visuomotorio è stata ampiamente associata all’attivazione del cervelletto sia a livello degli emisferi che del verme, con una attivazione prevalente degli emisferi in associazione ai movimenti della mano, e una attivazione prevalente del verme in associazione ai movimenti oculari (Miall et al., 2000). L’inattivazione funzionale del cervelletto laterale nella scimmia degrada
profondamente l’esecuzione di un compito di tracking visuomotorio (Miall et al.,1987). Analogamente, la lesione del cervelletto rende il paziente cerebellare incapace di
controllare la velocità e la traiettoria della mano mentre esegue un compito di tracking e induce tremore intenzionale al termine del movimento verso un bersaglio visivo.
Paradossalmente, la prestazione dei pazienti cerebellari migliora se vengono privati del feedback visivo, lasciando ipotizzare che il cervelletto sia responsabile non solo della elaborazione dei corretti comandi motori ma anche del controllo del movimento sotto la guida del feedback sensoriale, in particolare del feedback visivo (Holmes, 1917; Stein, 1986).
La gestione del feedback sensoriale rappresenta un problema di particolare difficoltà per il controllo motorio di un sistema biologico a causa dei lunghi tempi impiegati dalle informazioni sensoriali per raggiungere i centri deputati alla loro elaborazione. Infatti, il ritardo con cui il feedback sensoriale periferico, sia visivo che di altra modalità, agisce sul comando motorio induce instabilità del sistema, soprattutto quando sono richiesti movimenti rapidi (Miall et al., 1993). In questo caso, due strategie possono
teoricamente ridurre l’instabilità: l’intermittenza e la predizione. Nell’intermittenza, particolarmente evidente nel tracking manuale e nei movimenti saccadici oculari, il movimento è frazionato in sub-movimenti e ciascun sub-movimento è alternato a brevi periodi di immobilità necessari per aspettare l’arrivo del feedback e per programmare sulla base di questo i parametri del movimento successivo (Gross et al., 2002). E’ stato dimostrato che nel tracking sinusoidale a frequenza fissa, in cui la posizione del
bersaglio è facilmente prevedibile in ogni istante, l’ampiezza di ciascun sub-movimento è in relazione con la velocità con cui il bersaglio da inseguire si muove all’inizio del sub-movimento stesso (Miall et al., 1987). Verosimilmente, il cervelletto usa questa
informazione per prevedere la posizione del bersaglio quando il sub-movimento sarà compiuto e per programmare il movimento di conseguenza. Nel tracking non
sinusoidale o “pseudorandom”, in cui la posizione del bersaglio non è prevedibile, l’ampiezza dei sub-movimenti è invece prevalentemente in relazione con un parametro statico, l’errore posizionale, cioè la differenza fra la posizione del bersaglio e quella della mano in quel momento. Come nel caso del tracking “pseudorandom”, anche in caso di inattivazione cerebellare il soggetto perde la capacità di calcolare l’ampiezza di ciascun sub-movimento sulla base della velocità del target. Potendo fare affidamento soltanto sulla stima dell’errore posizionale, i suoi movimenti diventano ampi e veloci, con la tendenza ad oltrepassare il bersaglio (“overshoot”) e/o a muoversi iterativamente avanti e indietro rispetto ad esso (Miall et al., 1987).
Come accennato sopra, la predizione sensori-motoria è una strategia che il sistema motorio usa per ridurre l’instabilità del sistema legata al feedback sensoriale periferico. Nel controllo predittivo propriamente detto il feedback esterno è sostituito da un feedback interno fornito da un sistema neurale (“modello a forward”) capace di predire il feedback con ritardo trascurabile sulla base del comando motorio. Questa “previsione del feedback” può essere usata per la correzione dei comandi motori non appropriati assai precocemente rispetto al feedback reale, prevenendo l’instabilità del sistema. Oltre ad essere inviato ai centri di controllo (corteccia motoria) per la correzione del
movimento, l’output del modello interno a forward sarebbe anche opportunamente ritardato, per confrontarsi col feedback periferico proveniente in media 100-200 msec più tardi (non meno di 80-100 msec per l’input propriocettivo e 150-250 msec per l’input visivo). Nel caso in cui il segnale del feedback periferico sia prevedibile, e quindi già accuratamente previsto dal modello interno, esso sarebbe cancellato per non interferire con lo svolgimento del movimento; limitatamente alle informazioni non prevedibili, che nel caso di un compito appreso dovrebbero essere minime e dovute per lo più ad interferenze esterne, il feedback periferico, confrontandosi con il feedback previsto, potrebbe generare un segnale di errore che sarebbe usato per correggere il movimento. Questo sistema di controllo, in cui un modello a forward permette sia di prevedere il risultato del comando motorio che di gestire il feedback esterno, è noto in letteratura ingegneristica come “Smith predictor”. Una decina di anni fa è stato proposto che il cervelletto agisca come uno Smith predictor (Miall et al., 1993; Fig.I).
Il concetto di Smith predictor permette di spiegare perchè soggetti normali a cui è imposto sperimentalmente un feedback visivo ritardato presentino una esecuzione
imprecisa assai simile a quella dei pazienti cerebellari. Il ritardo del feedback esterno si traduce nell’ incapacità di gestirlo e di cancellarlo se prevedibile (Miall e Reckess, 2001). Lo “Smith predictor” consente di spiegare anche la degradazione dell’esecuzione del tracking nei pazienti cerebellari. Se il cervelletto è sede del modello a forward, è logico aspettarsi che la lesione cerebellare si traduca nell’incapacità sia di fornire un feedback interno che di gestire il feedback esterno. In questo modo lo Smith Predictor permette di spiegare la nota osservazione (vedi anche sopra) che i pazienti cerebellari con problemi motori di particolare gravità eseguono meglio il tracking in condizioni di “open loop”, cioè senza ausilio della vista basandosi su informazioni mnemoniche, piuttosto che di “closed loop”, cioè quando è fornito loro il feedback visivo.
Nonostante l’indiscutibile importanza del modello Smith predictor per aver introdotto il concetto di modello interno nel contesto del movimento biologico ed averlo localizzato nel cervelletto, negli ultimi anni questo modello ha mostrato una relativa inadeguatezza. Esperimenti di adattamento al ritardo del feedback visivo in soggetti normali che eseguivano un tracking manuale (Miall e Jackson, 2006) e altri tipi di studi di integrazione visuomotoria (Bhushan and Shadmehr, 1999; Flanagan et al., 2003) hanno mostrato che è necessario separare la predizione dal controllo. In altre parole, il maggior limite dello “Smith predictor” è il fatto che esso utilizza un singolo modello sia per la predizione delle conseguenze di un comando motorio che per il controllo dell’azione. L’idea corrente è che il movimento sotto controllo visivo necessiti di un modello a forward per la predizione delle conseguenze sensoriali di un comando motorio e di un modello inverso che predica il comando motorio appropriato per raggiungere il risultato desiderato.
La necessità di associare modelli a forward e modelli inversi e di incorporarvi il feedback è stata elaborata da almeno due gruppi di ricercatori. Nel modello di Bhushan e Shadmehr, il controllo motorio è realizzato dal modello inverso con la cooperazione del modello a forward che fornisce al modello inverso un segnale di errore sulla base della discrepanza fra stato predetto e stato desiderato (Bhushan e Shadmehr, 1999). Nel modello MOSAIC (Modular Selection And Identification Controller) il controllo motorio dipende da unità funzionali, dette moduli, ciascuna composta da un modello a forward e dal corrispondente modello inverso (Wolpert e Kawato, 1998). In questo modello (Fig.II C) si prende in considerazione il fatto che, a parità di atto motorio, il comando motorio deve tenere conto del contesto, per cui si deve postulare l’esistenza di più modelli interni inversi in grado di elaborare comandi motori diversi appropriati ai
diversi contesti. Il problema di selezionare il modello inverso più opportuno è risolto dal SNC generando per ogni modulo un segnale o “indice di responsabilità”. Prima dell’inizio del movimento l’indice di responsabilità stima sulla base del contesto la probabilità che ciascun modello ha di “soddisfare” il movimento voluto. A movimento iniziato, sulla base del confronto tra la predizione fornita dal modello a forward di ciascun modulo ed il feedback reale, l’indice di responsabilità valuta quanto ciascun modulo contribuisca al comportamento desiderato. Maggiore è l’indice di responsabilità, maggiore è la probabilità che il modulo simuli in maniera appropriata una certa azione in un dato contesto e partecipi al suo controllo.
Questo modello computazionale prevede anche che più moduli possano contribuire a simulare un’azione, ciascuno proporzionalmente al proprio indice di responsabilità nei confronti di quella azione, facendo sì che con un numero relativamente piccolo di moduli possa essere coperta l’intera rassegna di azioni sperimentabile nella vita.
E’ stato suggerito che il cervelletto sia una struttura ideale per generare e ritenere i modelli interni. Innanzi tutto è sede di fenomeni di plasticità sia a breve che a lungo termine che costituiscono il substrato per i processi di apprendimento motorio e di consolidamento delle memorie motorie (Ito, 2000 e 2001; Linden, 2003).
Secondariamente, il risultato di lesioni cerebellari osservate nell’uomo o indotte sperimentalmente nell’animale è compatibile con la presenza di modelli interni, in particolare modelli a forward, nella corteccia cerebellare (Wolpert et al, 1998; Kawato, 1999; Imamizu et al., 2000). Infine, studi di fMRI hanno evidenziato la comparsa di attivazione cerebellare durante l’apprendimento di un compito visuomotorio, che è che è stata interpretata come il correlato funzionale dell’acquisizione del modello interno del nuovo compito (Imamizu et al., 2000 e 2003). Più precisamente, mentre il soggetto imparava l’uso del mouse del PC in condizioni di rotazione della posizione del mouse sullo schermo, sono stati osservati due tipi di attivazione cerebellare: uno iniziale distribuito in una vasta area del cervelletto e proporzionale al segnale di errore, l’altro confinato in un’area più ristretta e posteriore del cervelletto e persistente anche dopo che l’apprendimento era concluso. Quest’ultimo tipo di attività è stato messo in relazione col l’acquisizione del modello interno del nuovo compito (Imamizu et al., 2000). Analogamente, è stato dimostrato che l’apprendimento di compiti diversi presuppone l’acquisizione di modelli cerebellari diversi e spazialmente segregati a livello della corteccia cerebellare, nonchè la possibilità di passare dall’uso dell’uno a quella dell’altro in maniera reciproca (Imamizu e al., 2003) .
Ovviamente nessuno studio di neuroimmagine può discriminare fra modelli diretti o a forward e modelli inversi. Anche se non mancano sostenitori della localizzazione intra-cerebellare dei modelli inversi (Kawato, 1999), i ricercatori hanno in genere proposto che il cervelletto sia sede di modelli diretti a causa della stretta associazione fra il modello a forward e l’ elaborazione dell’errore. L’errore, a sua volta, è stato da lungo tempo associato al cervelletto, fin da quando è risultato evidente che la proiezione olivo-cerebellare poteva veicolare un segnale di errore (Ito, 1928). Recentemente, studi di neuroimmagine hanno dimostrato che l’attività cerebellare correla con l’errore, sia che si tratti di un errore di esecuzione del compito visuomotorio (errore generato al soggetto), sia che si tratti di un errore di localizzazione del bersaglio, nel caso, per esempio, di un bersaglio che si sposta imprevedibilmente (errore imposto dall’ambiente; Diedrichsen et al., 2005).
Nell’insieme, le evidenze disponibili confermano ed estendono la relazione fra cervelletto ed errore motorio. E’ ovvio che se un segnale di errore è generato all’interno del cervelletto questo deve confrontarsi ed integrarsi col segnale di errore elaborato a livello corticale, principalmente a livello di PPC e ACC, come riportato precedentemente.
L’ipotesi che le attività cerebellare, frontale e parietale si integrino ai fini dell’elaborazione dell’errore, e in ultima analisi del controllo motorio on-line, necessita di almeno due premesse di ordine anatomo-fisiologico che un numero crescente di dati sperimentali sta via via confermando: a) che esistano “loop” cortico-cerebello-corticali che associano regioni specifiche della corteccia cerebrale con regioni specifiche del cervelletto, e b) che all’interno di ciascun loop l’informazione sia trasferita in maniera reciproca (Schmahmann e Pandya, 1997).
Per ciò che concerne le connessioni del cervelletto con la corteccia parietale, è stato dimostrato che esso riceve input dalla PPC via i nuclei pontini (Brodal e Bjaalie, 1997; Middleton e Strick, 1998a) e che proietta ad essa via il talamo (Clower et al., 2001). Per quanto riguarda le connessioni del cervelletto con la corteccia frontale, le evidenze sono ancora più impressionanti. Innanzitutto, nell’uomo la porzione ventrale del nucleo dentato, che proietta alla cortecce prefrontali, è più estesa di quella dorsale, che notoriamente proietta alla corteccia motoria (Matano, 2001). Analogamente, nell’ambito delle proiezioni cortico-pontine il maggior contingente di fibre afferenti al cervelletto è di origine prefrontale anziché motoria, come è invece tipico dei primati non umani (Ramnani, 2006). Studi di iniezione di traccianti retrogradi e anterogradi nella scimmia
hanno dato evidenza diretta che il cervelletto (in particolare Crus II) è reciprocamente connesso con la corteccia prefrontale (BA 9 e 46; Dum e Strick, 2003; Kelly e Strick, 2003). D’altra parte è noto che la corteccia prefrontale e la ACC sono reciprocamente interconnesse (Paus, 2001). In aggiunta, questi studi hanno precisato che le connessioni reciproche prefrontali sono anatomicamente segregate da quelle cerebello-motorie (che coinvolgono prevalentemente i lobuli cerebellari IV-VI). Questi dati confermano l’ipotesi di una organizzazione modulare delle connessioni cerebello-corticali per cui all’interno di ciascun loop l’attività può fluire da moduli cerebellari specifici a siti specifici della corteccia e viceversa (Middleton e Strick, 2001).
Infine, lo studio della coerenza del segnale RM nel cervelletto e in corteccia ha dimostrato l’esistenza di foci multipli di coerenza funzionale nel nucleo dentato e nelle aree prefrontali (BA 9, 10 e 46), con estensione alla corteccia cingolata (BA 24 e 32), nonché nelle cortecce parietale (BA 39 e 40) e occipitale (Allen et al., 2005), fornendo il substrato anatomo-funzionale all’ipotesi che il cervelletto influenzi l’operatività del circuito fronto-parietale nel processo di integrazione visuomotoria e di elaborazione dell’errore.
Fig.I Il modello dello Smith Predictor.
Il comando motorio (“motor command”) originatosi dalla corteccia motoria (“motor controller”) attiva il sistema motorio (“motor system”) che cambia di stato producendo il movimento e generando un cambiamento nella riafferenza sensoriale (“reafference”) che giunge al sistema nervoso centrale con un certo ritardo (“delay”).
Una copia del comando motorio (“efferency copy”) viene inviata ad un modello a forward, probabilmente contenuto nel cervelletto, il quale genera una stima dello stato motorio (“state estimate”). Tale stima è utilizzata all’interno di un loop a feedback negativo, caratterizzato da ritardo trascurabile e alto guadagno (indicato in figura dalle frecce circolari), capace di indirizzare il movimento verso lo stato desiderato (“desidered state”). Per permettere un utilizzo ottimale del feedback sensoriale lo Smith Predictor utilizza un altro modello (“forward output model”) che ritarda opportunamente la stima dello stato motorio consentendo il confronto con le riafferenze sensoriali. La discrepanza tra la predizione ritardata (generata dal “forward model” e dal “forward output model”) e il feedback reale è utilizzata per correggere il movimento. In ultima analisi lo Smith Predictor è formato da due loop: il primo è in grado di predire lo stato motorio, il secondo di trasformare e ritardare la stima per confrontarla con le riafferenze sensoriali. In tal modo qualunque errore interno ed esterno può essere corretto.
Fig.II Esempi di modelli interni.
A Modello inverso: i modelli inversi ricevano l’informazione sull’atto motorio desiderato e producono il comando motorio che permette di raggiungere la configurazione corporea appropriata.
B Modello a forward: i modelli a forward utilizzano la copia del comando motorio per stimare le conseguenze sensoriali del movimento.
C Modello MOSAIC: Il modello MOSAIC (Modular Selection And Identification Controller) consiste di una serie di unità funzionali, dette moduli (rappresentati in figura come n° fogli successivi), ciascuna composta da un modello inverso, dal corrispondente modello a forward e da un predittore di responsabilità (“Responsibility Predictor”). In particolare, prima dell’inizio del movimento, il predittore di responsabilità stima, sulla base del contesto (“contextual signal”), la probabilità che il modello inverso di ciascun modulo ha di “soddisfare” il movimento. A movimento iniziato, sulla base della copia efferente del comando motorio, il modello a forward genera una predizione del feedback sensoriale che si confronta con il feedback reale. Quanto più accurata è tale predizione tanto maggiore è la probabilità che il modello inverso contribuisca al movimento desiderato. I segnali generati dal sistema a forward e dal predittore di responsabilità vengono inviati ad un estimatore di responsabilità (“Responsability Estimator”) che “pesa” il contributo di ciascun modulo alla generazione del comando motorio a feedforward complessivo (“Feedforward motor command”) .
Scopo dello studio
Come riportato nell’introduzione, la corteccia frontale mediale, e in particolare la corteccia cingolata anteriore (ACC), provvede al monitoraggio on-line dell’azione segnalando la presenza di una discrepanza tra il risultato desiderato e il risultato ottenuto. Quest’ultimo è valutabile attraverso il feedback sensoriale o prevedibile ancor prima che sia disponibile il feedback, sulla base della copia efferente del comando motorio.
Il cervelletto, in quanto parte di un sistema di controllo dell’azione, contribuisce a generare un segnale di errore interno ogni qualvolta si rileva un conflitto tra predizione e esecuzione. In particolare è stato ipotizzato che il cervelletto contenga al suo interno modelli interni tra cui i modelli a forward (Wolpert et al., 1998), capaci di predire le conseguenze sensoriali di un azione basandosi sulla copia efferente del comando motorio.
Sulla base di queste evidenze sono state formulate le seguenti ipotesi:
- la corteccia frontale mediale è implicata nel rilevamento e/o elaborazione dell’errore in compiti di integrazione visuomotoria;.
- il cervelletto in virtù della sua capacità di predizione e di elaborazione dei segnali di errore, influenza direttamente e/o indirettamente l’operatività della corteccia frontale mediale.
Scopo dello studio è stato quindi duplice:
1) In primo luogo valutare le variazioni di attività e di connettività della ACC associate all’esecuzione di un compito visuomotorio e di correlarle all’errore commesso.
A tal fine, essendo il ritmo θ frontale mediale (fmθ) un indice di attivazione della ACC, sono state valutate le variazioni di potenza spettrale e di coerenza nella banda di frequenza θ di una sorgente di segnale EEG localizzata nella ACC durante un compito di tracking visuomotorio. E’ stato scelto questo tipo di compito in quanto a) la sua corretta esecuzione richiede il monitoraggio on-line dell’azione e/o dell’errore e quindi b) induce un elevato grado di cooperatività funzionale fra aree corticali diverse (Classen et al., 1998), in particolare fra quelle esecutive (frontali), associative (parietali), visive (occipitali) e motorie (corteccia motoria, supplementare motoria e premotoria).
Per escludere che gli effetti osservati dipendessero dalle componenti puramente motorie o puramente visive del compito, ai soggetti veniva fatto eseguire anche un compito puramente motorio, un compito puramente visivo e un compito visivo e motorio non richiedente integrazione visuomotoria e quindi il confronto continuo tra il risultato desiderato e risultato ottenuto.
2) Il secondo scopo dello studio è stato valutare se in presenza di deficit cerebellare fosse alterato il rilevamento e/o l’elaborazione dell’errore a livello della ACC.
A tal fine è stato utilizzato, in pazienti cerebellari, lo stesso disegno sperimentale e lo stesso approccio metodologico impiegato nella prima parte dello studio. Il compito di tracking visuomotorio è apparso anche in questo caso particolarmente indicato in quanto è noto che la sua corretta esecuzione dipenda dal controllo del cervelletto che fornisce il comando motorio appropriato (modello inverso) e predice le conseguenze sensoriali di tale comando (modello a forward) prima dell’arrivo del feedback sensoriale.
Materiali e metodi
Soggetti
Sono stati studiati 11 volontari sani con esame obiettivo neurologico negativo e anamnesi negativa per malattie psichiatriche e neurologiche (“ soggetti di controllo”, 5 maschi, età media 27±7anni, 2 mancini) e 8 soggetti con evidenza clinica di sindrome cerebellare e reperto radiologico di lesione prevalentemente unilaterale del cervelletto (“pazienti cerebellari”, 4 maschi, età media 41±16 anni).
Come riportato nella Tabella I al momento dello studio 7 pazienti presentavano una sindrome cerebellare acuta o subacuta con sindrome neurologica conclamata, mentre un paziente aveva una lesione cerebellare cronica con sindrome neurologica sfumata. Tutti i pazienti sono stati caratterizzati in termini di compromissione della funzione cerebellare in accordo con la International Cooperative Ataxia Rating Scale e in termini di localizzazione anatomica della lesione cerebellare sulla base degli esami TAC e MRI. Dal punto di vista eziologico la lesione cerebellare era di natura vascolare in 6 casi (emorragica in 3 e ischemica in altri 3 casi) e di natura neoplastica nei rimanenti 2 casi.
Lo studio è stato condotto in accordo con la “World Medical Association Declaration of Helsinki” e tutti i soggetti hanno dato consenso informato.
Disegno sperimentale
Entrambi i gruppi di soggetti, di controllo e pazienti cerebellari, sono stati sottoposti a due sessioni sperimentali in giornate non consecutive. I soggetti normali eseguivano i compiti con la mano dominante nel corso della prima giornata e con quella
controlaterale nel corso della seconda, mentre i pazienti adoperavano la mano ipsilaterale alla lesione cerebellare (“mano affetta”) nella prima giornata e quella controlaterale (“mano non affetta”) nella seconda, indipendentemente dalla mano dominante.
Ai soggetti veniva richiesta l’esecuzione di 4 tipi di compiti (task) di cui 3 motori: visuomotorio (VM), motorio propriamente detto (M) e visivo+motorio (V+M) e 1 puramente visivo (V), tutti della durata di 2 minuti, preceduti e seguiti da periodi di
stati registrati più periodi di VM, ciascuno della durata di 2 minuti alternati a periodi di rest sempre della durata di 2 minuti.
Durante i rest il soggetto fissava un punto luminoso immobile al centro dello schermo di un oscilloscopio posto di fronte a lui cercando di mantenere fissa la posizione dello sguardo e di evitare ammiccamenti (“blinking”).
Durante il task VM appariva sullo schermo dell’oscilloscopio un segnale bersaglio (target), cioè un cursore luminoso che si muoveva sinusoidalmente alla frequenza di 0.25 Hz con una ampiezza tale da sottintendere un angolo visivo di 4° nelle posizioni verticali massime dello sguardo corrispondenti al picco e al vallo della sinusoide.
Il task consisteva nell’inseguire il target riproducendone on-line il movimento sinusoidale ruotando fra il pollice e l’indice alternativamente in una direzione e nell’altra la manopola del potenziometro. Il soggetto controllava l’effetto del suo movimento sullo schermo dell’oscilloscopio dove era visualizzato come un secondo cursore luminoso e doveva graduare opportunamente l’ampiezza e la frequenza della rotazione effettuata in maniera da sovrapporre la sinusoide da lui prodotta a quella del target (Fig.1 A e B).
Nel task M il soggetto doveva produrre la stessa sinusoide che in VM ma privato sia della visione del target che di quella della sua esecuzione (performance), mentre in V+M doveva produrre la sinusoide ancora senza visione della sua performance ma con un input visivo distraente caratterizzato da una serie di stimoli luminosi che si modificavano in maniera random. Infine, nel task V il soggetto guardava la sinusoide sullo schermo dell’oscilloscopio senza effettuare alcun movimento. I task V, M e V+M precedevano sempre VM per evitare effetti secondari dovuti alla immaginazione del task di tracking.
Registrazione
Le singole tracce EEG sono state ottenute con registrazione monopolare (elettrodo di riferimento “linked earlobes”) da 19 elettrodi a coppetta montati su una cuffia elastica secondo il sistema internazionale 10-20 di posizionamento degli elettrodi. L’impedenza degli elettrodi era accuratamente mantenuta < 5 KΩ. I segnali erano amplificati e filtrati fra 0.03 e 70 Hz ed acquisiti alla frequenza di campionamento di 256 Hz con una scheda analogico-digitale a 12 bit. I segnali EEG sono stati convertiti off- line al sistema di riferimento “average linkage”.
Elettrodi a coppetta sono stati usati per la registrazione bipolare EMG dal primo muscolo interosseo dorsale ed dall’opponente del pollice di entrambi i lati per monitorare sia il movimento voluto che quello non voluto (per es. “mirror movements”). Elettrodi analoghi sono stati inoltre applicati in regione epicantale bilaterale per la registrazione bipolare dell’elettro-oculogramma (EOG) ed in regione precordiale per la registrazione dell’elettrocardiogramma (ECG), entrambi utili per identificare gli artefatti EEG dovuti all’attività elettrica e meccanica del cuore e/o a movimenti oculari ed ammiccamenti.
I segnali rappresentanti la sinusoide target e quella eseguita dal soggetto, sincronizzate con la registrazione EEG, sono stati entrambi registrati su PC (frequenza di campionamento 256 Hz con scheda analogica digitale a 12 bit) ed utilizzati per calcolare off- line l’errore posizionale.
Analisi
Analisi delle componenti indipendenti (IC). Dopo eliminazione degli artefatti maggiori,
i segnali EEG, EOG, EMG e ECG sono stati sottoposti all’analisi delle componenti indipendenti (Independent Component Analysis o ICA), una tecnica capace di separare le diverse sorgenti di segnale EEG comunemente sovrapposte a livello dei singoli elettrodi.
Lo scopo di questo approccio è stato duplice: 1) identificare e quindi scartare le componenti chiaramente artefattuali (dovute per es. ad attività muscolare, movimenti oculari e ammiccamenti), tanto più importanti nei soggetti affetti da patologie del sistema nervoso centrale; 2) ricavare sequenze di segnali EEG di interesse.
A questo scopo è stato applicato l’algoritmo dell’ICA messo a punto da Hyavarinen e Oja (1997) e implementato per MATLAB 5.x, versione 2.1 (gennaio 2001) nel pacchetto FastICA. La decomposizione è stata effettuata su un segnale a 22 canali (19 EEG, 1 EOG, 1 EMG e 1 ECG) della durata complessiva di 16 min (245760 punti) ottenuti concatenando periodi di rest e task (4 rest + 4 task) della durata di due minuti ciascuno.
Brevemente, questo algoritmo prevede che i dati EEG siano scritti come una matrice pxn, dove p è il numero dei canali EEG (22 canali) e n è l’estensione del vettore dati (245760 punti). Come output l’algoritmo fornisce una matrice di dimensione analoga a quella dell’input contenente il “time course” di attivazione delle singole componenti. Tali componenti sono ottenute mediante il calcolo della “unmixing matrix”, W, che
decompone i 22 segnali EEG originali nella somma di componenti massimamente indipendenti nel tempo e segregate nello spazio. Inoltre, le colonne della matrice inversa W-1 forniscono l’entità della proiezione di ciascuna componente su ciascun particolare elettrodo dello scalpo, per cui è possibile ricavare la proiezione topografica di ciascuna componente. Le attivazioni e le proiezioni topografiche consentono di distinguere le componenti artefattuali da quelle che rappresentano attività neurale.
I segnali EEG puliti possono essere riottenuti ri-proiettando e ri-mischiando sullo scalpo solo le componenti non artefattuali, moltiplicando la matrice contenente le loro attivazioni per la matrice W-1 .
La riproducibilità dell’analisi delle IC effettuate da FastICA è stata verificata con il programma Icasso per MATLAB di Himberg et al. (2004) che si basa sull’applicazione dell’algoritmo FastICA più volte (n=20) allo stesso segnale randomizzando ogni volta le condizioni iniziali. L’applicazione ripetuta dell’algoritmo crea stime ripetute delle singole componenti che vengono rappresentate da singoli punti nello spazio del segnale. Mediante l’analisi dei cluster le IC stimate sono raggruppate in cluster in accordo alla loro similarità. Se una IC ha una buona riproducibilità statistica stime ripetute di essa producono punti tutti prossimi alla IC “reale” e vicini tra di loro tali da segregare in cluster compatti ed isolati. Al contrario IC non affidabili corrispondono a cluster dispersi i cui elementi presentano similarità elevate con quelli di altri cluster. Per identificare i cluster più compatti ed isolati corrispondenti alle componenti statisticamente affidabili è stato introdotto un indice di qualità (Iq): esso è definito come la differenza fra la media delle similarità intra-cluster e la media delle similarità inter-cluster. Per un cluster ideale Iq è uguale ad 1 e tale valore decresce per cluster meno compatti ed isolati .
Poiché si assume che l’attività di una IC rifletta l’attività sincrona di una porzione relativamente ristretta di territorio corticale che può esser assimilata ad un singolo dipolo, la localizzazione di questo dipolo all’interno del cervello è stata calcolata sulla base della proiezione topografica della sua attività sullo scalpo. In termini quantitativi ciò corrisponde a trovare per una certa IC un singolo dipolo equivalente che spieghi la sua proiezione topografica con varianza residua inferiore al 20%. A tal fine è stato utilizzato il pacchetto DIPFIT del Software EEGLAB sviluppato da Oostenveld e Oostendorp (2002) per MATLAB impiegando il modello sferico standard di testa (quattro involucri: encefalo, meningi, spazio epidurale e teca cranica di raggio 71, 72, 79 e 85 mm e conducibilità 0.33, 1, 0.0042, 0.33 S/m, rispettivamente).
Le componenti i cui dipoli si localizzavano al di fuori del modello di testa standard o che presentavano varianza residua superiore al 20% sono state escluse dall’analisi.
Analisi spettrale. Nel presente studio l’ EEG “pulito” da artefatti è stato utilizzato per
calcolare il picco della frequenza α individuale (Individual Alpha Frequency o IAF) tipico di ogni soggetto così da determinare i limiti inferiore e superiore della banda theta (Klimesh, 1999). Lo IAF rappresenta la frequenza dominante (o picco) dell’ EEG del soggetto nel tradizionale range α 7.5-12.5 Hz. Usando lo IAF come “anchor point”, il θ è stato calcolato individualmente per ciascun soggetto come la banda di frequenza di 2 Hz tra IAF-4Hz e IAF-6 Hz.
L’analisi dello spettro di potenza e di coerenza è stata effettuata sulle componenti (“Indipendent components”, IC) isolate dall’ICA, selezionando fra quelle non artefattuali, le IC presenti in ogni soggetto che potevano ritenersi massimamente coinvolte nel task VM sulla base del “time corse” della loro attività e della loro localizzazione topografica.
Il calcolo della potenza spettrale è stato ottenuto utilizzando il metodo di Welch. Per ciascuna delle 7 IC selezionate è stato prima ottenuto un vettore dati della lunghezza di 30720 (256*120) per ogni periodo di task e di rest, che è stato segmentato in 119 epoche di 2 sec. parzialmente sovrapposte (“overlapping” 256, cioè del 50%). Ciascuna epoca è stata sottoposta ad analisi di Fourier usando una Fast Fourier Transform (FFT, numero di FFT 1024) previo “detrending” del segnale e applicazione di una finestra di Hanning (ampiezza 512 punti) per ridurre il “leakage spettrale”, ottenendo bins di 0.25 Hz.
La coerenza fra i segnali x e y appartenenti a coppie di IC indipendenti è stata calcolata utilizzando gli stessi vettori dati e gli stessi parametri di FFT (“linear detrending”, finestratura di Hanning, risoluzione spaziale di 0.25 Hz) usati per il calcolo della potenza spettrale, cioè vettori della lunghezza di 30720 punti segmentati in 119 epoche di 2 sec. parzialmente sovrapposte. E’ stato così possibile calcolare la coerenza secondo la formula
Cxy(f) = |Pxy(f) | 2 / Pxx(f)Pyy(f)
dove Pxx, Pyy e Pxy indicano gli autospettri e i cross-spettri dei segnali x e y. L’algoritmo
usato fornisce un numero reale compreso fra 0 e 1. Per ottenere valori di potenza spettrale e di coerenza riferiti alle singole bande è stata calcolata la media dei valori dei bins di frequenza che cadevano all’interno dei limiti definiti per ciascuna banda.
Infine, per ridurre la variabilità inter-soggetto ed intra-soggetto dei valori di coerenza assoluta è stato adottato il concetto di coerenza relativa, sottraendo la coerenza a riposo a quella durante il task in accordo con la seguente formula :
Cohdifference = Cohtask - Cohrest
Analogamente, anche la potenza spettrale è stata espressa come variazione percentuale del task rispetto al rest secondo la seguente formula:
%Powerchange= (Powertask – Powerrest) / Powerrest * 100
Analisi dell’errore. L’analisi dell’errore è stata calcolata in tutti gli 11 soggetti normali
per la mano dominante e in 7 per la mano non dominante, in 5 degli 8 pazienti cerebellari per la mano affetta e in 4 per la mano non affetta.
L’accuratezza del tracking durante il task VM è stata calcolata come errore posizionale (differenza tra la sinusoide tracciata dal soggetto e la sinusoide target) calcolato su epoche di 1 secondo, rettificato e integrato. Il valore ottenuto è stato quindi espresso come frazione del valore rettificato e integrato della sinusoide target ottenendo un indice di errore dove zero rappresenta la miglior performance e valori maggiori di 1 una performance scarsa.
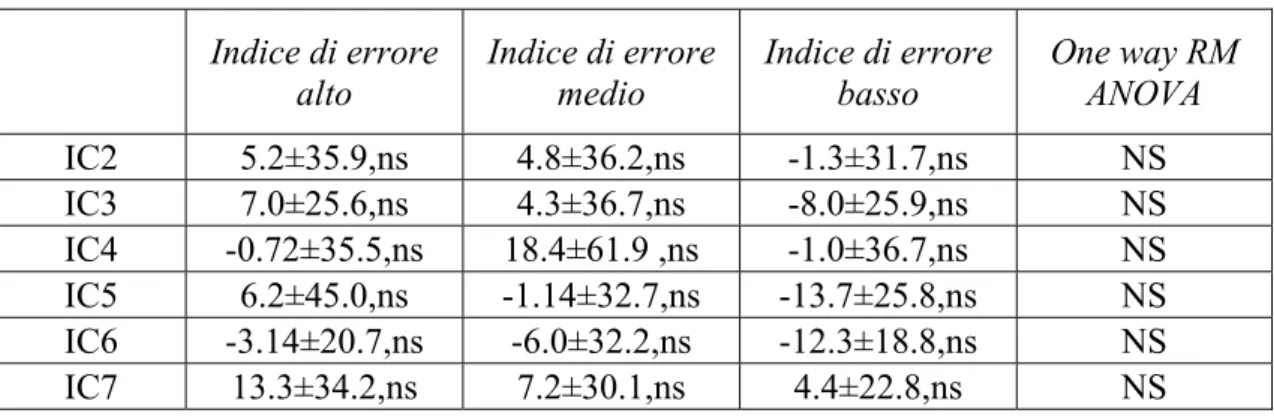
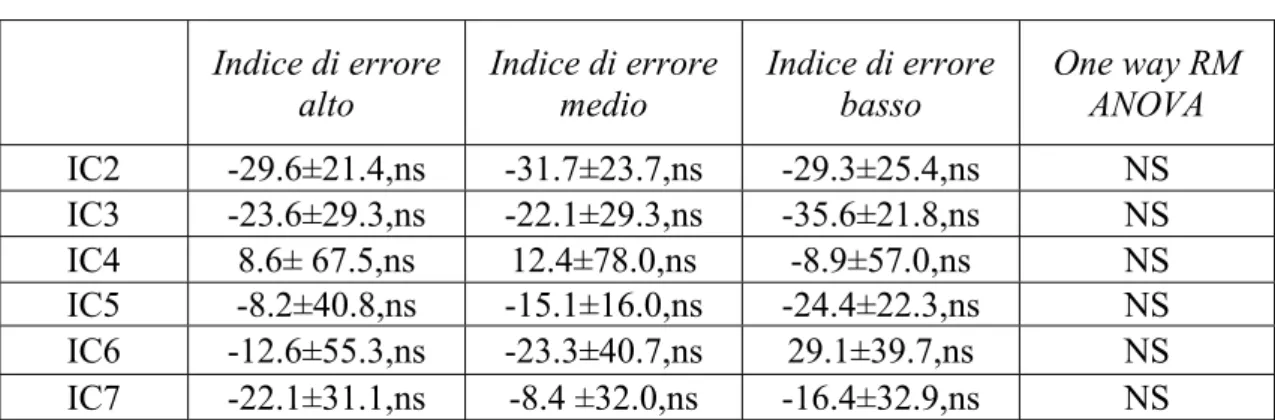
In base all’indice di errore le epoche di performance sono state ordinate in ordine crescente, in maniera da poter agevolmente selezionare le 50 epoche con l’indice più basso (performance accurata), le 50 con l’indice più alto (performance degradata) e 50 epoche con i valori di indice distribuiti intorno alla mediana.
Le epoche di EEG associate a indice di errore alto, medio e basso sono state utilizzate per studiare l’attività delle componenti in relazione con il livello alto, medio o basso di accuratezza della performance.
Analisi statistica. Prima di procedere all’analisi statistica la posizione delle IC a
localizzazione bilaterale simmetrica (IC2,IC3,IC4 e IC5) è stata invertita nei soggetti normali con mano dominante sinistra e nei pazienti cerebellari con lesione dell’emicervelletto sinistro in modo da normalizzare a destra sia i dati relativi alla mano dominante, per ciò che concerne i soggetti normali, sia quelli relativi alla mano ipsilaterale alla lesione (mano affetta) per i soggetti cerebellari.
In maniera analoga sono stati normalizzati a sinistra i dati per la mano non dominante dei soggetti normali e quelli per la mano controlaterale alla lesione (non affetta) per i pazienti cerebellari.Il confronto statistico fra soggetti normali e cerebellari è stato fatto
per tutti i dati normalizzati a destra e separatamente per tutti i dati normalizzati a sinistra.
Limitatamente all’analisi statistica i valori di potenza spettrale e di coerenza sono stati trasformati rispettivamente in logaritmo e arco tangente iperbolica, al fine di stabilizzare la varianza e aumentare la gaussianità dei dati.
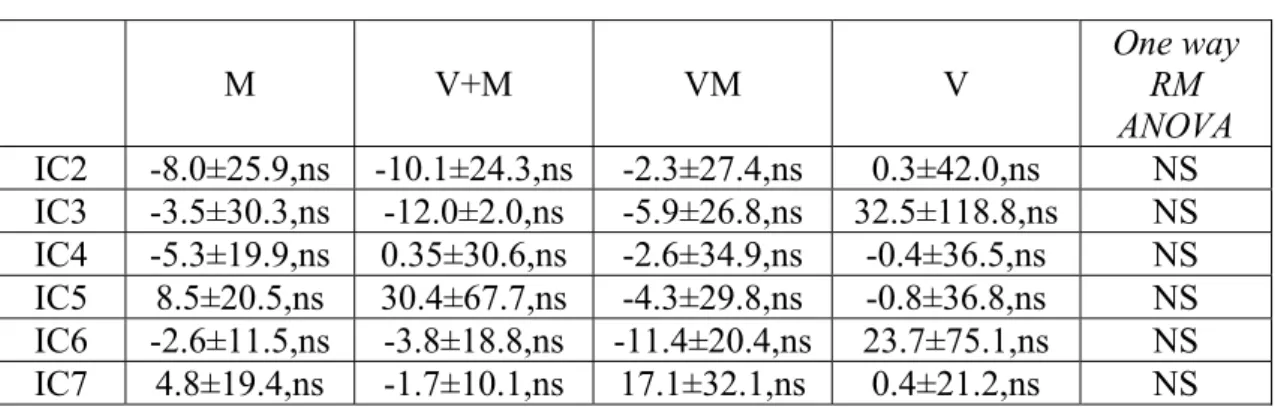
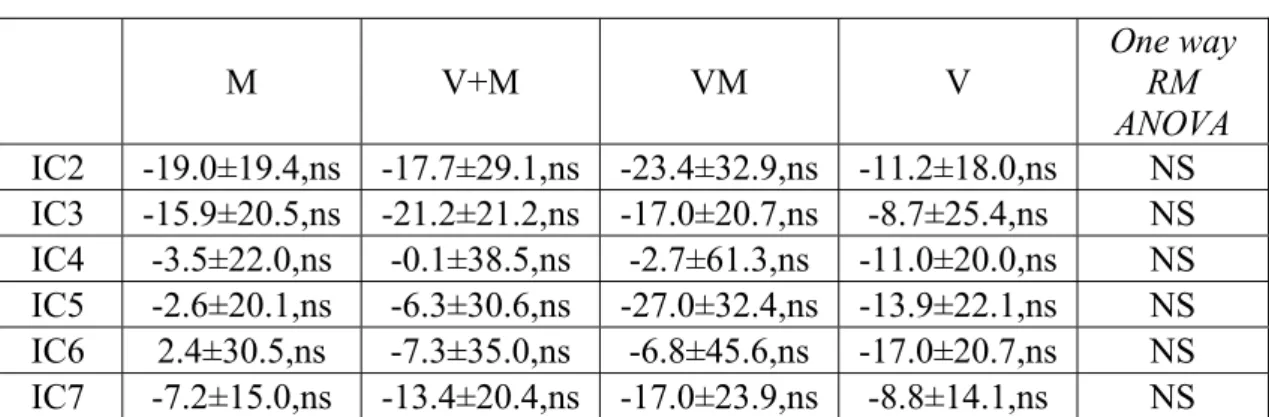
Le variazioni di potenza spettrale e di coerenza sono state calcolate rispettivamente per ogni IC e per ogni coppia di IC selezionate nell’ambito della banda di frequenza θ focalizzando l’attenzione su IC1 e IC6.
I dati sono poi stati espressi come valore medio ±DS, assumendo il livello di significatività per p< 0.05.
La significatività statistica delle variazioni di spettro e di coerenza associate al task rispetto al rest sono state valutate mediante il t-test per misure appaiate (paired t test), mentre quelle associate ai diversi task (V, M, V+M e VM ) mediante l’analisi della varianza ad una via per misure ripetute (one way RM ANOVA), utilizzando il test a posteriori Newman-Keuls se era rilevata una significatività. Il confronto fra soggetti normali e cerebellari è stato analizzato mediante analisi della varianza a due vie (two way ANOVA), cioè per condizione (normale e cerebellare) e per task (V, M, V+M e VM) utilizzando ancora, dove necessario, il test Newman-Keuls come test a posteriori. Analogamente, le variazioni di spettro e di coerenza in rapporto ai tre livelli di errore (indice di errore alto, medio, basso) sono state analizzate mediante one way RM ANOVA all’interno di ciascun gruppo, normale o cerebellare, mentre quelle fra i due gruppi mediante two way ANOVA (condizione x indice di errore, 2x3), utilizzando ancora in entrambi i casi il test Newman-Keuls come test a posteriori.
Risultati
Caratteristiche cinematiche dei task motori.
Caratteristiche cinematiche del task visuomotorio (VM). Nella Fig.2 sono illustrate le
caratteristiche cinematiche del task VM eseguito da un soggetto normale. Durante questo tipo di task appariva sullo schermo dell’oscilloscopio un segnale target (traccia blu), a cui il soggetto doveva sovrapporre un cursore (il cui movimento è indicato dalla traccia rossa) ruotando opportunamente la manopola del potenziometro. I soggetti normali eseguivano il task con relativa accuratezza come evidente soprattutto negli ultimi tre cicli del movimento riportato nella figura (Fig.2A). È in questa fase che si può osservare l’assenza di apprezzabili ritardi nel movimento del soggetto rispetto al target, indice della capacità di prevedere il movimento del target. Inoltre, una analisi più dettagliata della traccia del soggetto indicava che il movimento in apparenza continuo era composto di submovimenti che si ripetevano ad intervalli variabili da 100 a 300 ms (Fig.2 C). Per valutare quantitativamente l’accuratezza del tracking è stato calcolato per ciascun secondo di performance un indice di errore detto “errore posizionale” (vedi metodi): in particolare, un indice di errore uguale a zero si otteneva quando target e performance si sovrapponevano perfettamente, mentre valori maggiori di 1 indicavano una performance particolarmente degradata. La media dell’errore posizionale calcolato sulla base di tutti i valori di errore, uno per ciascun secondo di performance visuomotoria, è risultato 0.37±0.07 per la mano dominante di tutti i soggetti normali, indicando un discreto controllo della performance visuomotoria. Valori sovrapponibili si ottenevano per la mano non dominante (0.36±0.09). Per ciascun soggetto, l’errore posizionale calcolato durante il periodo iniziale della performance non differiva sostanzialmente da quello calcolato durante il periodo finale: tale fatto permetteva di escludere fenomeni quali l’apprendimento del task o l’affaticamento del soggetto. Rispetto ai soggetti normali i pazienti cerebellari eseguivano il task con accuratezza consistentemente minore. Più precisamente, essi riuscivano a seguire la frequenza del movimento imposta dal task ma non riuscivano a produrre movimenti di ampiezza appropriata come se mancassero della capacità di predire la posizione del target.
Ne deriva che nei 5 soggetti cerebellari l’errore posizionale medio, pari a 0.65±0.15 per la mano ipsilaterale alla lesione cerebellare, era significativamente diverso da quello ottenuto nei soggetti normali (unpaired t test, t=-5.261, p≤0.001, 5 pazienti cerebellari