Capitolo 2: Propriocezione e meccanocettori
Introduzione
La propriocezione è la capacità di conoscere la posizione e il movimento del proprio corpo (cinestesia), senza usare la vista. Le sensazioni che giungono al sistema nervoso centrale proveniendo dal sistema propriocettivo, di cui i meccanocettori sono l’ingresso, hanno primaria importanza per controllare i movimenti, manipolare oggetti, mantenere una corretta postura, sfruttando un circuito di retroazione del sistema nervoso centrale che interviene continuamente sui muscoli.
Nel corpo umano, il sistema propriocettivo è fortemente ridondante e complesso, difficilmente riproducibile allo stato attuale della tecnica.
Esistono tre tipi di meccanocettori nei muscoli e nelle articolazioni che contribuiscono ad informare sulla posizione e sulla velocità del movimento: fusi
muscolari (figura 2.1), organi tendinei di Golgi (figura 2.2), recettori localizzati nelle capsule articolari.
Nella pelle sono presenti meccanocettori cutanei e subcutanei (figura 2.3) sensibili a stimoli differenti: terminazioni di Ruffini, dischi di Merkel, corpuscoli
di Meissner, corpuscoli di Pacini, recettori follicolari, recettori di campo.
Perciò, è stato costruito un dispositivo che possa imitare alcune caratteristiche essenziali, come la capacità di rilevare allungamenti e pressioni sulla pelle.
2.1 Risposta elettrica generata dai meccanocettori
Le proprietà temporali di uno stimolo sono codificate dalla frequenza e dalla durata dell’attività dei neuroni sensoriali. Molti recettori rilevano la velocità alla quale lo stimolo aumenta o decresce d’intensità cambiando rapidamente la loro frequenza di scarica. Per esempio, quando una sonda tocca la pelle, il picco iniziale della frequenza di scarica è proporzionale sia alla velocità con la quale la pelle è indentata sia alla pressione applicata. Mentre la pressione è mantenuta costante, la frequenza di scarica rimane ad un livello proporzionale all’indentazione della pelle. La scarica cessa quando la sonda è allontanata.
Quindi, i neuroni rivelano la dinamica dello stimolo non solo quando scaricano, ma anche quando cessano di scaricare.
Esiste un fenomeno detto adattamento: a fronte di uno stimolo stazionario nello spazio e nel tempo la scarica del neurone tende ad attenuarsi dopo alcuni minuti. Questa è una caratteristica molto importante di tutti i recettori sensoriali: uno stimolo costante, non nocivo, tende ad essere ignorato dal sistema nervoso centrale, che altrimenti sarebbe sovraccaricato da una quantità immane di dati da elaborare.
2.1.1 Velocità di adattamento dei meccanocettori
I meccanocettori cutanei e subcutanei possono adattarsi allo stimolo rapidamente (FAR, Fast Adapting Receptors) presentando una risposta fasica, o lentamente (SAR, Slow Adapting Receptors) presentando una risposta tonica.
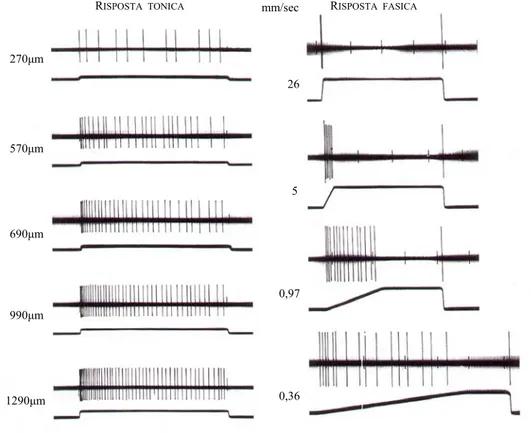
I SAR rispondono a stimoli prolungati, sono in grado di rilevarne l’intensità per diversi minuti generando una serie di potenziali d’azione della durata dello stimolo. In figura 2.4, a sinistra, è possibile osservare la risposta dei SAR a varie sollecitazioni costanti ma con diversi gradi di indentatura della pelle. Per ogni coppia, la prima traccia rappresenta i picchi dei potenziali d’azione che si generano, mentre la seconda indica la profondità (in micrometri) dell’indentatura. È evidente che al crescere della pressione il numero delle scariche dei potenziali d’azione cresce, concretando un incremento della velocità (della frequenza) di scarica, inoltre, la velocità di scarica è maggiore all’inizio e decresce mentre la pressione rimane costante, così i meccanocettori sentono quanto rapidamente la pressione è applicata sulla pelle. Le scariche cessano bruscamente quando lo stimolo è rimosso.
I FAR sono attivi solo all’inizio ed alla fine di uno stimolo. Essi cessano di scaricare quando l’intesità della sollecitazione diviene costante, mentre sono attivi solo in presenza di variazioni d’intensità: sono, quindi, sensibili alle derivate temporali dello stimolo (velocità e accelerazione). La risposta dei FAR dipende essenzialmente da due fattori: la prolungata depolarizzazione del potenziale del recettore inattiva il meccanismo di generazione nell’assone gangliare1; la forma del recettore si adatta allo stimolo costante, non generando più un segnale sufficiente a superare la soglia d’attivazione del potenziale d’azione.
In figura 2.4 a destra è illustrata la risposta dei FAR secondo diverse velocità di applicazione della sonda. Confrontando le coppie di grafici, è chiaro che i recettori non scaricano quando l’indentatura è costante. I grafici sono per indentature applicate con velocità (mm/sec) via via decrescenti, ma sempre a parità profondità finale: pressioni applicate lentamente generano una scarica lunga
1 I recettori non sono direttamente collegati al sistema nervoso centrale. Neuroni periferici, detti
neuroni gangliari, effettuano per mezzo di dendriti una giunzione sinaptica con i recettori; da questi neuroni si dipartono assoni che afferiscono al midollo spinale, ovvero sono presenti interneuroni tra il midollo e i neuroni gangliari.
nel tempo ed a bassa frequenza, indentature applicate rapidamente una breve scarica ad alta frequenza. Quindi, il movimento della sonda contro la pelle è caratterizzato sia dalla frequenza che dalla durata della scarica dei FAR.
mm/sec
RISPOSTA TONICA RISPOSTA FASICA
270µm 26 570µm 5 690µm 0,97 990µm 0,36 1290µm
Figura 2.4: Risposta dei SAR ( a sinistra) e dei FAR (a destra)
2.2.1 Fusi muscolari
Hanno una capsula che li racchiude e sono posti all’interno della parte carnosa dei muscoli. La loro funzione è di rilevare cambiamenti della lunghezza del muscolo che li ospita: variazioni in questo senso sono strettamente connesse
alla posizione angolare delle articolazioni, quindi il sistema nervoso centrale è in grado di valutare la posizione relativa delle vari e parti del corpo.
Ogni fuso ha tre componenti principali: un gruppo di fibre intrafusali con una porzione centrale non contrattile, terminazioni sensoriali mieliniche di grande diametro che efferiscono dalla parte centrale delle fibre intrafusali, terminazioni motorie mieliniche di piccolo diametro (motoneuroni γ) che innervano la parte contrattile delle fibre intrafusali. Esse sono disposte parallelamente al muscolo, quindi ne seguono gli stessi movimenti. Quando i fusi sono caricati meccanicamente, allungandosi, essi incrementano la frequenza di scarica dei potenziali d’azione; analogamente, ad una diminuzione della tensione muscolare corrisponde un decremento dell’attività nervosa.
I motoneuroni γ sono responsabili della contrazione della regione polare (parte contrattile) dei fusi, che, comunque, non contribuiscono alla forza espressa dalla contrazione del muscolo; l’attivazione dei motoneuroni γ serve a regolare la sensibilità dei fusi muscolari: contraendosi la regione polare, è incrementata la frequenza di scarica a parità di allungamento.
Le fibre intrafusali sono di due tipi: fibre con nucleo a sacco, dinamiche e statiche, e fibre con nucleo a catena. Simile suddivisione può essere fatta per le fibre nervose: le terminazioni sensoriali primarie (gruppo Ia)2, avvolte aspirale attorno alla regione centrale delle fibre intrafusali; le terminazioni sensoriali secondarie (gruppo II)3, che efferiscono dalle zone adiacenti a quella centrale delle fibre a sacco statiche e delle fibre a catena; i motoneuroni γ dinamici, afferenti4 alle fibre a sacco dinamiche; i motoneuroni γ statici che innervano le fibre a sacco statiche e le fibre a catena. La dualità della struttura è utile a distinguere diverse informazioni sulla posizione e sul movimento; la scarica
2 Le fibre sensoriali sono classificate secondo il loro diametro, in ordine decrescente dal gruppo I
al gruppo IV. Fibre di diametro maggiore conducono i potenziali d’azione più rapidamente.
3 Vedi nota 2.
4 In genere, i termini afferente ed efferente hanno come riferimento il sistema nervoso centrale, se
tonica5 delle terminazioni sensoriali primarie e secondarie fornisce dati sulla posizione stazionaria del muscolo. Inoltre, le terminazioni primarie sono altamente sensibili alla velocità d’allungamento ed a piccoli spostamenti, essendo, quindi, in grado di dare immediate indicazioni su mutamenti di lunghezza e permettendo rapide reazioni correttive: un classico esempio è il “senso dell’equilibrio”, che consente di mantenere la postura. L’incremento dell’attività dei motoneuroni γ dinamici aumenta la sensibilità dinamica delle terminazioni primarie, ma non ha influenza sulle terminazioni secondarie. L’incremento dell’attività dei motoneuroni γ statici aumenta l’attività delle terminazioni primarie e secondarie, abbassando la sensibilità dinamica delle terminazioni primarie e prevenendo l’inibizione dell’attività primaria quando il muscolo si accorcia. In questo modo il sistema nervoso centrale può indipendentemente regolare la sensibilità statica e dinamica delle fibre sensoriali nei fusi muscolari.
5 La scarica tonica, consistente in un segnale stazionario dei potenziali d’azione, può essere
prodotta sia da neuroni afferenti che efferenti. Ad esempio, un muscolo che deve essere mantenuto in uno stato non di riposo riceve una scarica tonica dai motoneuroni che gli afferiscono. Soggetti con lesioni relative a tali neuroni sono, ad esempio, in grado di muovere rapidamente l’occhio (movimento saccadico) verso l’alto, ma quasi immediatamente esso ritorna in posizione di riposo.
Fibra a Catena Nucleare Fibra a Sacco Nucleare Statica Fibra a Sacco Nucleare Dinamica II Ia Fibra Statica Fibra Dinamica Terminazioni Motorie γ Assoni Efferenti Assoni Afferenti Terminazioni Sensoriali Capsula Fibra Muscolare Intrafusale
Figura 2.1: Fuso Muscolare
2.2.2 Organi tendinei di Golgi
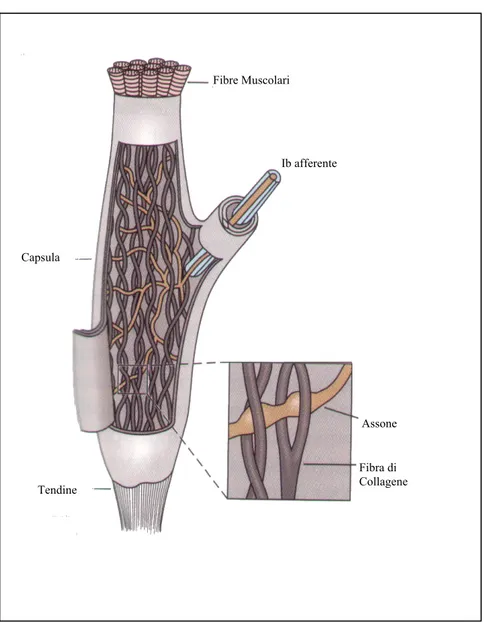
Sono localizzati alla giunzione tra le fibre muscolari e i tendini, essendo così in serie a gruppi di fibre muscolari, hanno forma allungata (1mm di lunghezza e 0,1mm di diametro) e presentano una capsula che li contiene (figura 2.2).
Ib afferente Tendine Capsula Fibre Muscolari Fibra di Collagene Assone
Figura 2.2: Organo di Golgi
Ogni organo è innervato da un singolo assone (gruppo Ib)6, che diviene privo di mielina all’interno della capsula e si divide in tante piccole terminazioni; queste terminazioni sono interposte tra le fibre intrecciate di collagene. Quando il muscolo si allunga, le fibre intrecciate di collagene si avvicinano tra loro comprimendo le terminazioni nervose che iniziano prontamente a scaricare; in
questo modo gli organi di Golgi sono sensibili non tanto all’allungamento, ma piuttosto alla tensione cui è sottoposto il muscolo.
2.3 Meccanocettori della pelle
Sono recettori molto simili tra loro, ma la diversa risposta alle sollecitazioni è dovuta alla diversa capsula che ha ciascuno di essi. Queste strutture sono disposte in maniera tale da essere deformate in modi particolari, allo scopo di rendere determinati recettori, ad esempio, sensibili alla pressione ma non all’allungamento. Studi fisiologici e istologici hanno identificato quattro tipi principali di meccanocettori cutanei: due sono situati negli strati superficiali della pelle e sentono le deformazioni delle sporgenze papillari dove sono localizzati; gli altri due tipi sono nei tessuti subcutanei e sentono deformazioni localizzate in un’area più ampia rispetto a quella fisicamente occupata da loro stessi.
Interfaccia Cute villosa Cute glabra Corpuscolo di Meissner Terminazioni libere Disco di Merkel Recettore Follicolare Corpuscoli di Pacini Epidermide Derma Fascio Nervoso Periferico
Figura 2.3: Disposizione schematica dei meccanocettori della pelle
2.3.1 Corpuscoli di Meissner
I corpuscoli di Meissner, appartenenti al primo tipo, rispondono rapidamente (FAR) ed essendo meccanicamente accoppiati al bordo delle sporgenze papillari reagiscono a stimoli molto ben localizzati. Possiedono una struttura globulare, che racchiude una pila di cellule epiteliali appiattite immersa in fluido extracellulare. Le terminazioni nervose efferenti sono innestate tra i vari strati della struttura.
2.3.2 Dischi di Merkel
Piccole cellule epiteliali intorno a terminazioni nervose costituiscono i
dischi di Merkel, localizzati generalmente in porzioni al centro delle sporgenze
papillari. La cellula di Merkel comprende una struttura semirigida che trasmette sforzi di compressione dalla pelle alla terminazione nervosa, evocando risposte notevoli ma adattandosi lentamente (SAR).
2.3.3 Corpuscoli di Pacini
I corpuscoli di Pacini, di tipo FAR, sono fisiologicamente simili a quelli di Meissner; rispondono a momentanee pressioni sulla pelle, ma non a stimoli stazionari a causa delle lamine di tessuto connettivo intorno alla terminazione nervosa. La grande capsula del recettore è collegata flessibilmente al derma, e ciò permette di rilevare vibrazioni verificantesi alcuni centimetri lontano dal corpuscolo.
2.3.4 Terminazioni di Ruffini
I meccanocettori di Ruffini sono di tipo a lenta risposta (SAR). Sono disposti nel tessuto subcutaneo intorno alle articolazioni, nel palmo e vicino alle unghie. Questi recettori sono sensibili all’allungamento e contribuiscono alla percezione della forma di oggetti.
2.3.5 Recettori follicolari e recettori di campo
I meccanocettori follicolari sono a risposta rapida (FAR) e si trovano nella pelle provvista di peli; sentono lo spostamento che dall’esterno si ripercuote nel follicolo. I meccanocettori di campo sono localizzati principalmente alle articolazioni delle dita, del polso, del gomito; rilevano l’allungamento e la frizione sulla pelle.