1
Sommario
1 INTRODUZIONE ... 3
1.1 Importanza salutistica del basilico ... 3
1.2 Metaboliti secondari del basilico ... 7
1.2.1 Radicali liberi e difese endogene ... 8
1.2.2 Gli antiossidanti ... 9
1.2.3 Lo stato fisiologico della pianta ... 12
1.3 Le interazioni dinamiche nel suolo ... 13
1.3.1 La simbiosi micorrizica ... 14
1.3.2 Effetti dei Funghi Micorrizici Arbuscolari sulla pianta ... 15
1.3.3 Microbiota associato ai funghi micorrizici arbuscolari ... 17
1.4. I batteri promotori della crescita della pianta (PGPB) ... 19
1.4.1 Effetti dei PGPB sulla pianta ... 21
1.5 Scopo della tesi ... 25
2. MATERIALI E METODI ... 27
2.1 Ceppi microbici ... 27
2.1.1.Trypticsoy agar (TSA) ... 27
2.1.2. Mezzo minimo con chitina colloidale... 28
2.1.3. Waksman agar ... 29
2.1.4. Nitrogen free mineral salts ... 29
2.2.1 Estrazione del DNA ... 31
2.2.2 Sequenziamento del 16S rDNA e analisi delle sequenze ... 32
2.3. Prova in vaso ... 33
2.3.1 Produzione dell’inoculo micorrizico ... 33
2.3.2 Potenziale dell’inoculo micorrizico ... 34
2.3.3 Preparazione dell’inoculo batterico... 34
2.3.4 Allestimento della prova sperimentale ... 35
2.3.5 Determinazione del peso fresco e del peso secco ... 35
2.3.6 Analisi microbiologiche ... 36
2
2.3.8 Analisi chimiche ... 41
2.3.9 Analisi molecolari ... 42
3. RISULTATI E DISCUSSIONE ... 48
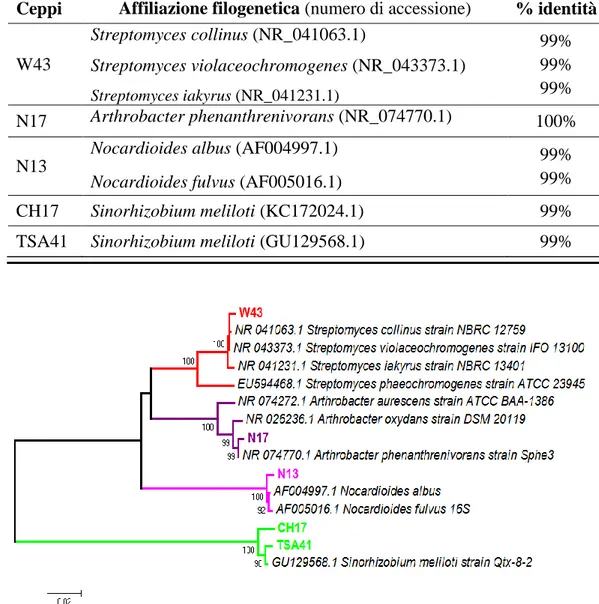
3.1 Identificazione dei ceppi batterici mediante analisi molecolari ... 48
3.2 Colonizzazione micorrizica di O. basilicum e potenziale di inoculo micorrizico ... 51
3.3 Determinazione del peso fresco e del peso secco ... 52
3.4 Analisi microbiologiche ... 53
3.5 Analisi biochimiche ... 54
3.5.1 Determinazione del contenuto in fenoli totali e della capacità antiossidante DPPH ... 54
3.5.2 Determinazione del contenuto in acido ascorbico ... 55
3.5.3 Determinazione della fluorescenza della clorofilla a ... 56
3.6 Analisi chimiche ... 58
3.6.1 Analisi del contenuto in azoto e fosforo... 58
3.7 Analisi molecolari ... 59
4 CONSIDERAZIONI CONCLUSIVE ... 67
3
1 INTRODUZIONE
1.1 Importanza salutistica del basilico
Il basilico (Ocimum basilicum, L.) è una pianta erbacea annuale appartenente alla famiglia delle Lamiaceae, comunemente coltivata come pianta aromatica. Per i suoi molteplici impieghi nelle preparazioni alimentari e farmaceutiche rappresenta una fonte di oli essenziali ricchi in composti fenolici, flavonoidi e antociani. Le foglie e le altre parti verdi del basilico sono tradizionalmente utilizzate come antispasmodico, sostanza aromatica, digestivo, e come agente tonico. Il basilico fresco viene utilizzato come ingrediente per vari piatti e per le preparazioni alimentari, soprattutto nella cucina mediterranea. Grazie alla sua attività antimicrobica e insetticida e al piacevole aroma, l’olio essenziale di basilico è ampiamente usato nelle industrie alimentari, farmaceutiche, cosmetiche e in aromaterapia.
Dal punto di vista botanico O. basilicum è una pianta aromatica annuale, longidiurna, con altezza compresa tra i 50 e i 70 cm, i cui fusti sono eretti e tendono a ramificare, se privati dell’apice, e provvisti di foglie opposte, picciolate, ovate-lanceolate, con margini interi. L’apparato radicale si presenta fascicolato e con alto potere penetrante nel terreno. I fiori, riuniti in verticilli, sono portati in posizione apicale con corolle da bianco a rosato, da cui si raccoglie il seme. La pianta è una specie originaria delle aree calde asiatiche ed africane, anche se adesso è diffusa in tutto il mondo, in particolare in ambienti con estate calda. Il basilico è molto esigente in fatto di terreno, clima e cure colturali, pertanto richiede temperature di vegetazione > 25°C e terreni ricchi in sostanza organica e ben drenati. Date tali caratteristiche, la sua coltivazione estensiva si riscontra in Italia, Francia, Egitto, Indonesia, Marocco, in numerosi Stati degli Stati Uniti, in Grecia, in Israele ed in alcuni Paesi dell'America Latina (Tucker et al., 2009). In Italia la coltivazione in serra ha subito negli anni notevoli variazioni (dati ISTAT), rimanendo in ogni caso uno tra i paesi a più alta produzione, grazie al suo rilevante impatto economico (dati ISMEA 2013) (Tabella 1 e 2).
4
Tabella 1: Produzione agricola totale di basilico per anno (quintali)
Anno 2005 2006 2007 2008 2009 2010 2011
Basilico in serra 495 545 1.337 369 938 231 355
Tabella 2: ISMEA–Osservatorio Economico del settore delle piante officinali. Giugno 2013: Rilevanza
economica del basilico
Nome Botanico Ocimum basilicum L
Habitat Eurasia
Area produzione cosmopolita
colt/spont coltivazione
Impiego aromi, alimenti, integratori, cosmetici, farmaci,
mangimi
Parti utilizzate foglia, sommità, olio, estratti
Utilizzo annuo (Kg) 57.840
Valore medio unitario (Euro/Kg) 1,63
Valore (Euro) 94.279
Il basilico è un’erba officinale aromatica di particolare interesse in quanto può essere utilizzata per la produzione di materie prime o preparati contenenti sostanze fitochimiche con significative capacità antiossidanti, benefiche per la salute (Exarchou et al., 2002). L’interesse nei confronti di questa pianta non è motivato dalle sole caratteristiche nutraceutiche, ma anche dall’ampio e versatile utilizzo in tutto il mondo e dalla numerosa varietà di cultivar disponibili e coltivabili in serra, consentendone la manipolazione controllata delle condizioni di crescita (Toussaint, 2007).
Il crescente interesse dei consumatori verso il consumo di “functional food”, per la prevenzione e il mantenimento della salute nei confronti delle malattie, è un aspetto importante per lo studio di fitochimici facilmente fruibili dai vegetali (Copetta et al., 2006). La dieta certamente gioca un ruolo importante nelle principali malattie umane: le malattie cardiovascolari, il cancro, l'ipertensione e l'obesità. Le stime accettate dalla comunità scientifica dimostrano che almeno un terzo dei casi di cancro e la metà dei casi di malattie arteriose e ipertensione sono
5 legati al tipo di dieta adottata (Block et al., 1999). Ciò evidenzia come il dilemma alimentare non appartenga esclusivamente al terzo mondo, ma, in senso contrario, sia presente anche nei paesi sviluppati e in via di sviluppo (Sökmen et al., 2004). La maggior parte delle malattie è dovuta alla generazione di radicali liberi, prodotti naturalmente dal metabolismo aerobio, che tendono ad aumentare a seguito di eventi patologici come l'infiammazione. In generale, gli antiossidanti fermano il processo delle reazioni a catena dei radicali liberi, aiutando a prevenire lo sviluppo di eventi patologici. L'interesse verso le vitamine C ed E, e verso i fenoli è dovuto alle loro proprietà antiossidanti (Sgherri et al., 2010) che proteggono le funzioni biologiche sia nel metabolismo vegetale che animale. L’impiego degli antiossidanti, in campo alimentare, per la prevenzione delle malattie legate allo stile di vita, ha un ruolo fondamentale anche per gli additivi alimentari che prevengono e ritardano il deterioramento alimentare.
Negli anni passati gli antiossidanti sintetici sono stati ampiamente utilizzati in molti alimenti per ritardare i cambiamenti indesiderati dovuti all'ossidazione e, tra questi, i più utilizzati erano il butilidrossianisolo (BHA) ed il butilidrossitoluolo (BHT), molecole fenoliche sintetiche liposolubili che agiscono come antiossidanti primari particolarmente attivi. La messa in commercio di prodotti aggiunti con BHA e BHT risale al 1958, periodo in cui erano stati considerati innocui e sicuri per la salute. Tuttavia, è stata successivamente rilevata la loro potenziale tossicità e cancerogenicità, soprattutto ad elevate concentrazioni. Pertanto l'uso alimentare di butilidrossianisolo (BHA) e butilidrossitoluolo (BHT) è stato limitato. In questo contesto si colloca l’intensificata ricerca di nuove fonti di antiossidanti d’origine vegetale (Sgherri et al., 2010).
Gli antiossidanti sono composti che possono ritardare o inibire l'ossidazione dei lipidi o di altre molecole, inibendo l'innesco o la propagazione di reazioni ossidative a catena (Marotti et al., 2010). L'effetto antiossidante è dovuto principalmente ai componenti fenolici, e a diverse vitamine, come la C e la E. L'attività antiossidante, garantita dai composti fenolici, è principalmente dovuta alle proprietà redox di tali molecole, che possono giocare un ruolo importante nel neutralizzare i radicali liberi (Phuong et al., 2008).
6 Negli ultimi anni, anche gli oli essenziali e gli estratti a base di erbe hanno suscitato un grande interesse scientifico in quanto potenziali fonti di antiossidanti naturali e composti biologicamente attivi (Bozin et al., 2006). L’attività antimicrobica e antiossidante degli oli essenziali delle piante aromatiche è stata sfruttata per svariate applicazioni farmaceutiche nella medicina alternativa. In particolare gli oli essenziali sono stati usati nelle terapie naturali per il trattamento delle malattie infettive, al fine di presentarli come valida alternativa alla medicina tradizionale (Bozin et al., 2006).
Molte piante aromatiche appartenenti alla famiglia delle Lamiaceae, come la salvia, l’origano, e il timo, mostrano una forte attività antiossidante (Hirasa & Takemasa, 1998). Il basilico appartiene a questa famiglia, con circa 60 varietà e cultivar, distinguibili per l’aspetto e l’aroma (Simon et al., 1999), ma eguagliati dalla presenza in composti fenolici con spiccata attività antiossidante (Nakatani, 1997). Gli studi di Galangal et al. (2005) hanno evidenziato l’efficacia antiossidante del basilico in diverse condizioni di temperatura e pH. I risultati hanno mostrato che l’attività antiossidante è ottimale a pH neutro e decrescente in ambienti acidi, mentre, anche a temperature elevate, l’attività antiossidante risulta stabile. Il lavoro di Abdullah Ijaz Hussain et al. (2008) ha mostrato che il ruolo di scavenger del basilico nei confronti delle specie reattive dell’ossigeno (ROS), è influenzato dalle condizioni ambientali e stagionali. Inoltre, nella stessa ricerca, gli autori hanno valutato l’attività antimicrobica dell’olio essenziale di basilico nei confronti di batteri patogeni come Staphylococcus aureus, Escherichia coli, Bacillus subtilis, Pasteurella multocida e di funghi patogeni come Aspergillus niger, Mucor mucedo, Fusarium solani, Botryodiplodia theobrome e Rhizopus solani, stabilendone anche la minima concentrazione inibente. Tutti i dosaggi antimicrobici per ciascun microrganismo hanno mostrato che tutti i patogeni, fungini e batterici, sono stati influenzati dall’attività antimicrobica del basilico. L’interesse nei confronti delle attività inibitorie del basilico su certe infezioni croniche ha condotto alla sperimentazione di due composti antiossidanti: l’acido rosmarinico e l’α-tocoferolo sull’HIV (virus dell’immunodeficienza acquisita), dimostrandone la sua efficacia (Yamasaki et al., 1998).
7 In aggiunta all’attività antiossidante esplicata dai composti fenolici, tra i metaboliti secondari ad azione fitochimica del basilico si evidenzia anche la presenza di antiossidanti vitaminici tra cui la vitamina C, la cui concentrazione fogliare è influenzata da molti fattori, tra cui il suolo, l’irrigazione e le condizioni climatiche (Velioglu et al., 1998).
I metaboliti ad attività fitochimica vengono prodotti dalle piante durante il loro sviluppo e accrescimento in risposta ad una serie di stimoli esterni, quali la difesa contro i microrganismi, contro gli insetti o erbivori in genere (Crozier et al., 2006; Herms et al., 1992), la disponibilità di nutrienti (Herms et al., 1992), l’esposizione a radiazioni ultraviolette (Rozema et al., 1997) e nelle interazioni allelopatiche (Mann et al., 1987). Tuttavia, poiché le risposte delle piante a tali stimoli sono varie, aumentare il valore nutraceutico non è facile. In particolare, la disponibilità di macronutrienti essenziali, durante la crescita delle piante, ha un significato fondamentale nell’influenzare l'accumulo di fitochimici (Phuong et al., 2008). L’apporto di questi componenti limitanti può essere agevolato ed implementato dall’impiego di bioinoculi a basso impatto ambientale ma ad alto rendimento nella crescita della pianta. I funghi micorrizici arbuscolari (AMF) e i batteri della micorrizosfera sono tra i principali agenti in grado di promuovere la crescita radicale e quindi l’assrobimento degli elementi nutritivi, promuovendo così la produzione di sostanze fitochimiche antiossidanti, benefiche per la salute (Artusson et al., 2006).
1.2 Metaboliti secondari del basilico
La fitochimica studia la grande varietà di sostanze organiche scoperte che, accumulandosi nelle piante, ne assicurano la presenza di sostanze bioattive. Inoltre, tale scienza sta definendo anche la struttura di questi composti, la loro biosintesi, il metabolismo, la distribuzione naturale e le attività biologiche (Harborne et al., 1998). Un posto importante tra queste è occupato dalle piante aromatiche, tra cui il basilico, il cui valore nutrizionale è associato alla presenza di oli essenziali, miscele complesse di composti volatili, dominate da mono e sesquiterpeni. Oltre agli oli essenziali, il basilico è caratterizzato anche dalla
8 presenza di composti fenolici, soprattutto cumarine e fenilpropanoidi, che hanno dimostrato di possedere molteplici attività farmacologiche. Gli studi, per determinare l'attività antiossidante del basilico, sono principalmente concentrati sulle attività antiossidanti di estratti grezzi, utilizzando come solventi metanolo, acetone, o acqua.
1.2.1 Radicali liberi e difese endogene
In ogni organismo aerobio avvengono processi biochimici che consumano ossigeno per la generazione di energia. Dal processo di utilizzazione dell’ossigeno (ossidazione), si producono tuttavia dei prodotti potenzialmente dannosi: i radicali liberi dell’ossigeno. Dal punto di vista chimico i radicali liberi sono molecole particolarmente instabili, in quanto presentano uno o più elettroni spaiati nell’orbitale più esterno. Questo induce i radicali liberi alla ricerca del proprio equilibrio chimico tramite acquisizione degli elettroni mancanti da altre molecole che, di conseguenza, perdendo un elettrone diventano a loro volta instabili e vanno quindi alla ricerca della stabilità cercando l’elettrone da altre molecole, innescando così un meccanismo di instabilità a catena.
Tra le molecole altamente reattive rientrano le ROS come l’anione superossido, il radicale idrossilico, il radicale perossidico e l’ossido d’azoto, nonché specie non radicaliche, come il perossido d’idrogeno, l’ossigeno singoletto, l’acido ipocloroso e l’ozono; tra questi, indubbiamente, il radicale idrossilico è la ROS più reattiva e dannosa nei sistemi biologici. Tale radicale è generato dalla reazione dello ione ferroso (Fe++) con il perossido d’idrogeno. In questo processo, noto come reazione di Fenton, la quantità di radicale prodotto è direttamente proporzionale alla concentrazione di ferro o rame (Mittler, 2002).
E’ indubbio che negli organismi animali esista un sistema endogeno in grado di abbattere le ROS che normalmente si formano durante i processi biochimici. Tuttavia, i sistemi endogeni, seppure attivi, possono non fornire una sufficiente protezione contro le ROS quando queste vengono prodotte in eccesso; in questi casi assumono particolare rilievo gli antiossidanti introdotti con la dieta. Oltre a danneggiare le cellule e a causare gravi malattie, le ROS sono responsabili dei processi di ossidazione negli alimenti, tra cui il principale è l’ossidazione lipidica.
9 Tale processo riduce la shelf-life dei cibi, provocando alterazioni organolettiche e riduzioni del valore nutrizionale.
1.2.2 Gli antiossidanti
I composti antiossidanti appartengono ad una classe molto eterogenea, con differenti strutture e caratteristiche chimiche (idrofilici o idrofobici), diversa distribuzione in natura, livello di concentrazione in sistemi biologici, siti di azione, efficacia contro le specie ossidanti, nonché presenza di altre azioni biologiche oltre quella antiossidante. La capacità antiossidante complessiva di frutta e ortaggi può essere determinata per mezzo di diversi saggi che utilizzano diverse sostanze radicaliche come l’acido 2,2'-azino-bis(3-etilbenzotiazolin-6-sulfonico) (ABTS), il 2,2-difenil-1-picrilidrazile (DPPH) e saggi che indirettamente determinano la capacità di scavenger delle specie radicaliche come la capacità di ridurre il ioni ferro (FRAP), la valutazione della concentrazione minima Cidal (MCC) e la capacità dell'ossigeno di assorbire i radicali (ORAC) (Kovtun et al., 2000).
Alcune classi di antiossidanti ben conosciute, come i carotenoidi, la vitamina C, i tocoferoli e i polifenoli meritano particolare attenzione. Questo non solo perché sono quelle maggiormente presenti negli alimenti, in particolare in frutta e verdura, ma anche perché sono assorbiti e metabolizzati in maniera differente ed hanno funzioni diverse, tutte molto importanti per garantire la salute umana (Nakatani, 1997). Javanmardi et al., (2003) hanno misurato la capacità antiossidante con il metodo ABTS, usando come standard il potere antiossidante dell’acido 6-idrossi-2,5,7,8- tetrametilcroman-2-carbossilico (Trolox), evidenziando la presenza di attività antiossidante nel basilico iraniano.
È molto importante valutare oltre al contenuto di composti antiossidanti, anche la loro biodisponibilità. Esistono in bibliografia diverse definizioni di biodisponibilità, tuttavia secondo Porrini e Riso (2008), è da intendersi come quella frazione dei nutrienti o composti ingeriti che raggiungono il sistema circolatorio o lo specifico sito, in cui essi esercitano la loro azione biologica (Balasundram et al., 2005).
10 Il contenuto nelle piante di composti bioattivi in genere e antiossidanti nello specifico è influenzato da vari fattori interni ed esterni alla pianta, sia nel periodo pre- che post-raccolta. Uno di questi fattori è sicuramente la disponibilità dei vari elementi nutritivi. Fanasca et al., (2006) hanno notato, ad esempio, che nel pomodoro un elevato quantitativo di potassio nella soluzione nutritiva incrementa nei frutti il contenuto di composti antiossidanti, specialmente di licopene.
La maggior parte degli antiossidanti che si conoscono appartengono al regno vegetale. Molte piante sintetizzano questi composti per soddisfare varie necessità fisiologiche: la difesa contro parassiti, agenti tossici, condizioni ambientali inadeguate, raggi ultravioletti e l'attrazione degli impollinatori (gli antociani e i flavonoidi sono responsabili della colorazione di fiori e foglie). Queste funzioni avvengono a spese del metabolismo primario delle piante (accrescimento e riproduzione) in quanto parte degli assimilati sono destinati allo svolgimento delle suddette attività (metabolismo secondario). Una funzione particolarmente importante del metabolismo secondario è la protezione da stress ossidativi; infatti, le piante, essendo organismi sessili e alla base della catena alimentare, hanno un attivo metabolismo secondario e sono quindi più ricche in sostanze antiossidanti (Allen et al., 2005).
Gli antiossidanti naturali, ampiamente presenti in alimenti quali frutti e ortaggi, ricadono essenzialmente in tre grandi gruppi: le vitamine, i composti fenolici e i carotenoidi. L’acido ascorbico ed i fenoli sono tra i più noti antiossidanti idrofili, i carotenoidi tra quelli lipofili. Anche alcuni minerali (selenio, rame e zinco) svolgono indirettamente funzioni antiossidanti, poiché partecipano alla regolazione di enzimi coinvolti nel meccanismo di difesa antiossidante (Balasundram et al., 2005).
Alcuni autori hanno evidenziato, invece, la forte influenza che può avere il genotipo; in merito a questo aspetto Balasundram et al. (2005) hanno evidenziato che, mentre certi composti antiossidanti, quale l’acido ascorbico, sono ubiquitari, altri sono specifici di certe famiglie o addirittura specie.
11
1.2.2.1 Gli antiossidanti vitaminici
La vitamina C, comunemente nota come acido ascorbico (γ-lattone dell’acido 2-chetogulonico) è caratterizzata da uno spiccato potere riducente a causa del gruppo enediolico. Il suo prodotto di ossidazione è l’acido L-deidroascorbico. Le piante, ed alcuni organismi animali, sono in grado di sintetizzarla a partire dai glucidi, mentre l’uomo deve introdurla necessariamente con gli alimenti in quanto non è in grado di effettuare il processo di sintesi. Nelle piante è coinvolta, insieme alla glutatione-perossidasi, nella detossificazione del perossido d’idrogeno. La vitamina C è il più potente antiossidante idrosolubile presente nel plasma sanguigno e agisce anche come rigeneratore della vitamina E e di specie cationiche radicaliche di carotenoidi. E’ noto, ad esempio, l’effetto sinergico delle vitamine E, C e del β-carotene come scavengers nei confronti di specie reattive dell’azoto.
Dal punto di vista delle funzioni biologiche, l’acido L-ascorbico agisce da cofattore enzimatico, da “radical scavenger”, nonché da donatore/accettore nel trasporto di elettroni nelle membrane plasmatiche.
1.2.2.2 Gli antiossidanti fenolici
Sono una classe di composti organici naturali caratterizzati dalla presenza di almeno un anello aromatico, mono o pluri-sostituito da gruppi ossidrilici. Una classificazione generale delle principali classi di composti fenolici prende in considerazione lo scheletro carbonioso che costituisce l’asse fondamentale per la differenziazione strutturale. L’attività antiossidante dei polifenoli è legata all’elevato potenziale redox, che consente loro di agire come agenti riducenti, donatori di idrogeno e “spegnitori” (quencher) dell’ossigeno singoletto. In aggiunta, possono agire come chelanti dei metalli coinvolti nella generazione delle specie radicaliche. La loro attività antiossidante è considerata molto più elevata di quella delle vitamine.
Condizione fondamentale, affinché sia esplicata l’attività antiossidante dei composti fenolici, è la formazione di radicali fenolici stabili, attraverso la delocalizzazione elettronica sulle strutture aromatiche ed alifatiche.
12 I composti fenolici principali del basilico sono l’acido rosmarinico, l’acido vanillinico, l’acido cumarinico, l’idrossibenzoacido, l’acido siringico, l’acido ferulico e l’acido caffeico (Javanmardi et al., 2002).
1.2.3 Lo stato fisiologico della pianta
Il processo biochimico di organicazione della CO2, che porta alla formazione di
carboidrati, quindi composti organici, è noto come fotosintesi ed avviene grazie alla conversione dell’energia luminosa in energia chimica. La fotosintesi è il processo attraverso cui l’energia solare viene assorbita dai pigmenti fotosintetici e convertita in energia chimica e, nello specifico, nel legame chimico delle molecole di ATP e NADPH. Tali coenzimi, prodotti dal trasporto elettronico cloroplastico, sono essenziali per i processi biochimici del ciclo di Calvin, attraverso cui la CO2 viene ridotta a carboidrati, ma sono essenziali anche per
l’organicazione di altri elementi oltre al carbonio, come l’azoto e lo zolfo.
Il processo ha inizio con l’assorbimento dell’energia luminosa da parte dei pigmenti fotosintetici nelle foglie, in particolare le clorofille. A seguito dell’assorbimento dell’energia luminosa, la molecole della clorofilla si eccita raggiungendo una condizione chimicamente instabile. Essa deve quindi dissipare l’energia in eccesso per tornare allo stato fondamentale. I meccanismi di dissipazione sono alla base del processo fotosintetico (trasferimento di energia di risonanza e foto-ossidazione). Esistono tuttavia altri due meccanismi deputati alla dissipazione dell’energia di eccitazione: il calore e la fluorescenza.
La fluorescenza della clorofilla a consiste nella ri-emissione di un fotone ad un lunghezza d’onda più elevata (in quanto a minore contenuto energetico) e rappresenta una via di dissipazione della energia antitetica ai processi fotochimici anche se concomitante a questi. La misurazione della fluorescenza della clorofilla a fornisce informazioni sulla funzionalità fotochimica e sulla dissipazione dell’energia sotto forma di calore, propri del processo fotosintetico (Daley et al., 1989; Raschke et al., 1990; Mott et al., 1993; Genty e Meyer, 1994; Bro et al., 1995; Siebke e Weis, 1995).
La determinazione della fluorescenza della clorofilla a rappresenta un efficace e affidabile, ma non invasivo metodo utilizzabile per la valutazione dei processi
13 connessi all’assorbimento e conversione dell’energia luminosa sulle membrane del cloroplasto. A tal fine è stato ed è tutt’oggi ampiamente utilizzato per la valutazione degli effetti di stress biotici ed abiotici nelle piante. Analisi sull’emissione della fluorescenza della clorofilla a sono state effettuate da Shimon Meir et al. (1997) su foglie intatte della pianta di basilico (Ocimum basilicum L) per valutarne l’entità dei danni causati dal freddo.
1.3 Le interazioni dinamiche nel suolo
Il suolo, secondo la definizione proposta dalla Soil Conservation Society of America (SCSA) (1986) è un corpo naturale costituito da particelle minerali ed organiche che si forma dall'alterazione fisica e chimico-fisica della roccia e dalla trasformazione biologica e biochimica dei residui organici. Capace di sostenere la vita delle piante, è caratterizzato da un’atmosfera interna, da una flora e da una fauna determinate e da una particolare economia dell'acqua. Rappresenta il mezzo di interazione dinamica tra atmosfera, litosfera, idrosfera e biosfera (SCSA, 1986). Le interazioni che avvengono in questo ambiente ne determinano le sue proprietà come mezzo per la crescita e il sostentamento della flora e della fauna microbica, facendone un parametro chiave per la qualità e la produzione agricola. Al fine di tutelare questo habitat, numerosi ricercatori hanno posto l’attenzione sull’impiego di prodotti chimici in agricoltura che rischiano di depauperare la ricchezza biologica del suolo e, quindi, influenzare negativamente l’ambiente e la produzione delle colture (Artusson et al., 2006). In ambito biotecnologico la tutela della biodiversità del suolo è un aspetto importante in quanto, dalle interazioni del microbiota terricolo, dipende la produzione sostenibile (Barea et al., 2004).
La porzione del suolo circoscritta all’apparato radicale è identificata come rizosfera. A livello di questa porzione di terreno, nella zona dell’interfaccia con le radici, i microrganismi e i costituenti del suolo interagiscono con esse per sviluppare un ambiente dinamico e responsabile della crescita e della salute delle piante negli agroecosistemi (Lynch, 1990).
I funghi micorrizici arbuscolari (AMF) rappresentano un importante gruppo di microrganismi benefici della rizosfera, in grado di condizionare in modo
14 significativo la crescita delle piante. La porzione di suolo adiacente alle radici delle piante e agli AMF, in cui i funghi AM e i batteri del terreno interagiscono, prende il nome di micorrizosfera (Linderman, 1988). Nella maggior parte dei casi le interazioni sono sinergiche e l’insieme di queste cooperazioni rizotrofiche e dei loro conseguenti effetti sulle proprietà del suolo e, quindi, sulla crescita della pianta, possono avere importanti implicazioni ecologiche ed agricole (Miransari, 2011).
1.3.1 La simbiosi micorrizica
Il termine micorriza deriva dalla combinazione dei due termini greci “funghi” e “radici” e descrive un’ampia varietà di associazioni tra questi due elementi. Le micorrize sono presenti in molti ambienti e il loro successo ecologico riflette un alto grado di diversità nelle capacità genetiche e fisiologiche degli endofiti fungini. Le micorrize si suddividono in endo ed ecto-micorrize; quest’ultime formano un mantello ifale attorno alle punte delle radici degli alberi forestali cambiandone la morfologia mentre le endomicorrize cioè arbuscolari (AM), micorrize ericoidi, e orchidee, hanno ife che penetrano nelle cellule della radice, in modo da stabilire una simbiosi intracellulare a prescindere dalla pianta ospite. Alcuni aspetti del processo di colonizzazione sono comuni a tutti i funghi micorrizici, sia che essi formino ectomicorrize (ECM), che endomicorrize arbuscolari (AM). In quanto funghi del suolo, i simbionti fungini sviluppano nella rizosfera grazie ai propaguli (spore e ife rizomorfe), che costituiscono il ponte tra terreno e radici. Tali ponti sono responsabili della cosiddetta “wood wide web”, rete di connessioni tra piante diverse che formano un percorso per il trasporto orizzontale dei nutrienti (Simard et al., 1997). Molte piante eterotrofe ottengono carbonio organico collegandosi al livello di questa rete di connessione, per mezzo delle micorrize endofitiche (Selosse et al., 2009).
Il rapporto che si instaura tra la pianta e il partener micorrizico è una simbiosi mutualistica, in cui entrambi i partner traggono vantaggio. Generalmente si sostiene che i funghi micorrizici migliorino l'assorbimento dei nutrienti vegetali grazie all'esplorazione ifale della rizosfera, e, in cambio, ricevono dalle piante
15 carboidrati, essenziali per il completamento del ciclo vitale (Thompson et al., 2009).
I principali vantaggi che le piante traggono dall’interazione con gli AMF riguardano il miglioramento dell’assorbimento dei nutrienti, la protezione contro gli stress ambientali e colturali e il miglioramento della struttura del suolo. Tali benefici sono dovuti allo sviluppo della rete ifale, che si estende dalle radici micorrizate al suolo circostante. Inoltre i funghi AM sono dei fattori fondamentali del funzionamento e della biodiversità degli ecosistemi, in quanto ogni cambiamento nella loro popolazione comporta cambiamenti nella composizione della comunità vegetale in termini di sopravvivenza, competizione e diversità floristica (Giovannetti, 2001). Tuttavia diverse pratiche colturali influenzano negativamente il ruolo degli AMF in agricoltura sostenibile, tra queste sono compresi i trattamenti chimici, i biocidi, la monocoltura e l’impiego di piante non in grado di sviluppare simbiosi micorrizica. Pertanto la selezione di opprtune pratiche agricole, che fanno uso di biofertilizzanti favoriscono la maggiore efficienza della simbiosi micorrizica in campo.
1.3.2 Effetti dei Funghi Micorrizici Arbuscolari sulla
pianta
Gli aspetti pratici e funzionali dell’impiego in agricoltura dei funghi AM hanno da molto tempo richiamato l’attenzione sull’uso di tali funghi come biofertilizzanti. I funghi AM sono infatti noti per la loro capacità di valorizzare l’apporto idrico e l'assorbimento di nutrienti limitanti alle piante (Baslam et al., 2013). Quest’ultimo aspetto viene esplicato aumentando la solubilità e, quindi, l'assorbimento di diversi macro e micronutrienti alla pianta. Ad esempio il fosforo è tra i nutrienti più importanti e limitanti, il suo assorbimento viene notevolmente incrementato nella pianta attraverso l’azione delle reti ifali extraradicali che si estendono ben al di là della zona di esaurimento che si forma intorno alle radici (Giovannetti e Avio, 2002). I funghi AM possono anche indurre resistenza sistemica nella pianta, come uno strumento necessario alla pianta per resistere agli agenti patogeni (Liu et al., 2007). Inoltre, i funghi AM possono anche alleviare gli effetti negativi di
16 stress del suolo sulla crescita delle piante e sulla produzione delle colture, rivestendo un ruolo significativo come bioprotettori (Xavier e Boyetchko, 2002). Non solo la pianta trae beneficio dalla simbiosi con gli AMF ma, agendo sui costituenti minerali e sull’approvvigionamento idrico, anche il suolo ne trae vantaggio, migliorando la propria struttura (Azcón-Aguilar e Barea, 1997).
Questi fattori indicano l'importanza dei funghi AM nel settore biotecnologico in ambito agro-industriale. Tale impiego in agricoltura è stato comparato con altre tecniche per la promozione della crescita e la qualità nutrizionale della lattuga (Baslam et al., 2013). Secondo questo studio l’impiego dei funghi micorrizici arbuscolari pare essere una valida alternativa rispetto all’utilizzo di altre tecniche. Inoltre, la medesima ricerca, ha evidenziato che gli effetti benefici dei funghi micorrizici sulla produzione e qualità sono altamente dipendenti da altri fattori, come il tipo di lattuga, la specie degli AMF, le condizioni colturali, il regime idrico applicato, la stagione e l’apporto di CO2 in serra (Baslam et al., 2013a). Un’ulteriore studio sulla lattuga ha mostrato che l’applicazione del fungo micorrizico G. fasciculatum era in grado di favorire la crescita della cultivar, in quanto la simbiosi apportava fattori nutrizionali e acqua, grazie alle ife esterne. Lo studio ha mostrato che, non solo la crescita viene promossa ma anche altre risorse vengono movimentate dalla macchina fotosintetica, ciò è stato avvalorato da un maggiore apporto in clorofilla e/o in carotenoidi (Baslam et al., 2013b).
Il conseguente incremento di biomassa, dovuto al maggiore apporto di nutrienti e acqua, in relazione all’impiego di AMF, è stato riportato anche in piante di Artemisia annua. Lo studio (Rapparini et al., 2008) ha infatti evidenziato che le piante bioinoculate con AMF della specie Glomus presentavano apparato radicale più lungo, un maggior numero di foglie e un più elevato peso secco.
In condizioni di stress salino, è stato evidenziato, da misurazioni dell’efficienza fotosintetica, che la salinità riduce drasticamente la crescita e la fluorescenza delle cultivar di basilico. In queste condizioni l’inoculo micorrizico ha evidenziato di ridurre i sintomi dello stress salino, data la ripresa della crescita e i valori della fluorescenza della clorofilla (Zuccarini et al., 2008). Altri studi hanno osservato come la colonizzazione micorrizica possa migliorare le prestazioni fisiologiche
17 della pianta stressata, portando a rendimenti più elevati e a maggiore qualità (Rosendahl e Rosendahl, 1991).
Come la maggior parte delle piante, anche le piante officinali beneficiano della simbiosi mutalistica con AMF, migliorando la produzione e l'accumulo di importanti principi attivi come terpeni, fenoli e alcaloidi; ottimizzando la composizione di diversi principi attivi e, infine, migliorando la qualità dei materiali a base di erbe (Yan Zeng, 2013). Attualmente, un’importante problema nei sistemi di coltivazione delle piante officinali è la qualità instabile dei prodotti. In questo contesto le micorrize arbuscolari (AM), influenzando il metabolismo secondario e la produzione di principi attivi delle piante medicinali, si presentano come un possibile mezzo per influenzare la qualità delle erbe a scopo fitochimico (Yan Zeng et al., 2013). Recenti studi hanno mostrato che piante del genere Origanum in simbiosi con G. mossae mostravano, oltre ad un incremento nel contenuto in fosforo, anche un aumento nel contenuto in olio essenziale (Toussaint et al., 2007) (Khaosaad et al., 2006). Anche la simbiosi AMF-basilico potrebbe essere impiegata come nuovo approccio per incrementare e mantenere la qualità degli oli volatili del basilico (Prasad et al., 2011).
L’interesse nei confronti della simbiosi basilico/AMF è dettato, non solo dall’incremento in biomassa e fitochimici che essa comporta, ma anche dalla protezione della cultivar da agenti di stress biotico, come può essere Fusarium oxysporum. Infatti è stato ipotizzato che G. mosseae potrebbe conferire un effetto bioprotettivo contro Fusarium oxysporum, riducendo la mortalità della pianta del 20%; questa ipotesi è stata avvalorata da un aumento delle foglie e dall’incremento in acido rosmarinico e caffeico in piante infette (Toussaint et al., 2008).
1.3.3 Microbiota associato ai funghi micorrizici arbuscolari
Le spore e le ife dei funghi micorrizici arbuscolari formano un micro habitat unico e particolarmente adatto alla colonizzazione di molte specie batteriche. Dal 1970, alcuni microrganismi, soprattutto batteri, sono stati osservati in stretta associazione con i funghi AM. Questi batteri vivono sulla superficie o tra le spore e le ife del fungo, nella parete sporale, ed anche all'interno del citoplasma fungino
18 (Bianciotto et al., 1996). Molti di questi batteri hanno effetti benefici sulle attività degli AMF, compresa la promozione della germinazione delle spore fungine, della crescita ifale, della colonizzazione delle radici, e anche la formazione di nuove spore fertili. Questi effetti suggeriscono l’importanza di questi batteri nel ciclo di vita del loro ospite fungino (Long et al., 2008).
Alcuni autori, usando tecniche colturali, hanno isolato diverse comunità batteriche che vivono in stretta associazione con le spore di AMF. Tra questi, Bharadwaj et al., 2008 hanno isolato 385 batteri dalla superficie delle spore di Glomus intraradices e G. mosseae. Questi batteri sono stati testati per la loro capacità di inibire la crescita in vitro di Rhizoctonia solani e per la loro produzione di siderofori. I generi più abbondanti che sono stati identificati sono Arthrobacter e Pseudomonas; la loro presenza è inoltre influenzata dalla specie di fungo AM e, in una certa misura, anche dalla cultivar considerata.
Un ulteriore lavoro di identificazione, coltura dipendente, è stato effettuato da Cruz et al. (2007). In questo studio sono stati isolati e identificati dai protoplasti di Gigaspora margarita due ceppi batterici Janthinobacterium lividum (KCIGM01) e Paenibacillus polymyxa (KCIGM04), con effetti antagonisti su numerosi funghi patogeni quali: Rosellimia necatrix, Pythium ultimum, F. oxysporum e R. solani. In particolare J. lividum ha mostrato maggiore attività bioprotettiva, mentre P. polymyxa ha evidenziato una maggiore solubilizzazione del fosforo.
Allo scopo di ottenere maggiori informazioni sulla diversità batterica associata alle spore sono stati utilizzati studi molecolari coltura indipendenti. In particolare, gli studi di Roesti et al. (2005), usando la tecnica della reazione a catena della polimerasi (PCR) ed elettroforesi su gel in gradiente denaturante (DGGE), hanno analizzato il microbiota batterico strettamente associato alle spore di Gigaspora margarita. I risultati hanno evidenziato la presenza di batteri appartenenti ai phyla Proteobacteria e Actinobacteria. Infatti, i membri degli Actinobacteria sono stati di solito trovati in associazione con AMF, sebbene non siano stati rilevati in associazione con le spore di Glomus geosporum e Glomus constrictum mediante analisi DGGE.
19 Tra gli Actinobacteria il genere Streptomyces è stato identificato in altri lavori (Tylka et al., 1991; Mugnier e Mosse, 1987). Da questi autori è stato mostrato che la germinazione delle spore è favorita dalla presenza degli Actinobacteria.
La diversità di batteri strettamente associati agli AMF, può dipendere dal fungo stesso, a causa della competizione per i nutrienti del suolo (Christensen e Jakobsen, 1993).
Tra i batteri della micorrizosfera sono presenti particolari batteri che agiscono promuovendo l'attività e lo sviluppo dei funghi AM, i Mycorrhiza Helper Bacteria (MHB) (Frey-Klett et al., 2007). Essi sono di solito fungo-specifici ma non pianta-specifici (Rillig et al., 2005) e possono influenzare la germinazione delle spore, agendo sulla parete (de Boer et al., 2005), producendo sostanze stimolanti come la CO2, o modificando l’assorbimento del fosforo da parte del fungo
(Bonfante et al., 2002). Tali batteri, date le loro attività biofertilizzanti sulle piante a interesse alimentare, rispondono all’esigenza di tecniche colturali maggiormente rispettose per l’ambiente, in grado di promuovere la micorrizzazione controllata o bioremediation microbica.
1.4. I batteri promotori della crescita della pianta (PGPB)
Con il termine Plant Growth Promoting Bacteria (PGPB) vengono considerati i batteri che migliorano la salute, la crescita delle piante e la fertilità del suolo in quanto liberi nel terreno, ma altamente competitivi nella colonizzazione della rizosfera. Il termine PGPB fu utilizzato per la prima volta da Klopper et al. nel 1980, nonostante fossero già noti dal 1978 dal medesimo ricercatore, e comprende batteri che vivono a vita libera nel suolo, indipendentemente da cellule vegetali, e batteri localizzati in strutture specializzate, internamente alle cellule vegetali. Il genere più noto appartenente a questo gruppo di batteri rizosferici è Rhizobium, ma sono presenti anche altri generi noti per le loro attività di promotori della crescita delle piante e utilizzanti come biofertilizzanti, quali: Azotobacter, Azospirillum, Bacillus, Klebsiella sp., Rhizobium, Bradyrhizobium, Azorhizobium, Sinorhizobium, Mesorhizobium e Paenibacillus (Hayat et al., 2010). L’attuale intensificata ricerca su i PGPB ha condotto alla realizzazione di una nuova
20 disciplina, che ha cambiato i tradizionali concetti di base della fisiologia vegetale e dell’ecologia microbica (Soalno et al., 2008).
I batteri benefici della rizosfera PGPB, colonizzanti le radici, sono distinguibili da tre caratteristiche intrinseche:
- devono avere la capacità di effettuare la colonizzazione radicale;
- devono sopravvivere e moltiplicarsi in microhabitat associati alla superficie delle radici, in continua concorrenza con il microbiota nativo, almeno per il tempo necessario a esprimere le loro attività di promozione delle piante.
- devono avere la capacità di promuovere la crescita della pianta (Barea et al., 2006).
I PGPB, caratterizzati da questi aspetti, sono atti a svolgere molti processi ecosistemici, come quelli coinvolti nel controllo biologico dei patogeni della pianta, nel ciclo dei nutrienti e/o nella stabilizzazione della pianta.
Studi basati su tecniche molecolari hanno stimato più di 4.000 specie microbiche per grammo di suolo rizosferico (Montesinos, 2003). Tra questi, gli attinobatteri filamentosi sono considerati come una delle più importanti comunità della rizosfera, essendo capaci di influenzare lo sviluppo della pianta e proteggere le radici dai fitopatogeni. Gli essudati vegetali, come gli aminoacidi e gli zuccheri, forniscono una ricca fonte di energia e di nutrienti per i batteri, con conseguente aumento delle popolazioni microbiche nella porzione micorrizosferica.
I cambiamenti strutturali della comunità microbica micorrizosferica sono stati indagati con l'applicazione della reazione a catena della polimerasi (PCR) ed elettroforesi su gel in gradiente denaturante (DGGE). Le indagini hanno mostrato numerose differenze, dipendenti dalle diverse associazioni instauratesi tra batteri e piante diverse (Herschkovitz et al., 2005).
La colonizzazione rizosferica da parte dei batteri è considerata come un passo cruciale nell'applicazione di microrganismi a scopi benefici come la biofertilizzazione, la fitostimolazione, il biocontrollo e la fitodepurazione (Compant et al., 2010).
21
1.4.1 Effetti dei PGPB sulla pianta
Le indagini sui meccanismi di promozione della crescita delle piante, attuati dai ceppi di batteri PGP, hanno mostrato che i meccanismi generali includono: fissazione dell’azoto, abbassamento dei livelli di etilene, produzione di siderofori e fitormoni, induzione alla resistenza contro i patogeni, solubilizzazione dei nutrienti, promozione dell’attività benefica dei funghi micorrizici arbuscolari e diminuzione della tossicità degli inquinanti (Glick et al., 1999).
Generalmente la promozione della crescita può avvenire per mezzo di due meccanismi: diretti e indiretti (Glick et al., 1999). La stimolazione diretta include la fissazione biologica dell’azoto, la produzione di fitormoni come le auxine, le citochinine e le giberelline, la solubilizzazione minerale del fosforo e del ferro, la produzione di siderofori ed enzimi e l’induzione della resistenza sistemica. La stimolazione indiretta, invece, è legata alla lotta biologica, comprende perciò la produzione di antibiotici, il processo di chelazione del ferro disponibile nella rizosfera, la sintesi di enzimi extracellulari in grado di idrolizzare la parete fungina e, in generale tutte le strategie attuate per colonizzare le nicchie della rizosfera (Zahir et al., 2004).
In base al loro meccanismo d’azione, i PGPB possono essere classificati con tre funzioni generali: biofertilizzanti, fitostimolanti e biopesticidi (Martinez Viveros et al., 2010). I generi Allorhizobium, Azorhizobium, Bradyrhizobium, Rhizobium e Sinorhizobium sono descritti come i generi PGPB a maggiore attività biofertilizzante (Vassey 2003). Questi microrganismi operano migliorando lo stato nutrizionale della pianta ospite, agendo quindi da biofertilizzanti e incrementando le associazioni benefiche del suolo nella pianta.
Più di uno, tra i meccanismi fitostimolanti, biofetilizzanti e biopesticidi può essere svolto anche da un solo PGPB, per migliorare la crescita delle piante. Infatti, le prove sperimentali suggeriscono che la stimolazione della crescita è il risultato di molteplici meccanismi che possono essere contemporaneamente attivati (Martinez Viveros et al., 2010). L’evidenza di queste molteplici attività è stata riportata in molti lavori di ricerca. Walker et al. (2012) hanno mostrato le diverse attività di due generi con spiccate attività PGP: Azospirillum e Pseudomonas. Questi hanno dato risultati diversi se usati singolarmente o in combinazione, ma in entrambi i
22 casi sono stati osservati cambiamenti qualitativi e quantitativi nella produzione di metaboliti secondari. Tuttavia, a causa della loro capacità di conferire alle specie vegetali effetti benefici, la colonizzazione efficiente della pianta è una fase della massima importanza. La maggior parte dei batteri associati ai vegetali deriva dall'ambiente terreno. Essi possono migrare verso la rizosfera e successivamente al rhizoplane dei loro ospiti prima che siano in grado di mostrare effetti benefici. Alcuni batteri, gli endofiti, colonizzanti il rhizoplane, possono anche penetrare nelle radici delle piante, e alcuni ceppi possono muoversi verso le parti aeree delle piante, con una densità batterica minore rispetto alla rizosfera. La comprensione dei processi di colonizzazione è importante per prevedere come i batteri interagiscono con le piante e se sono capaci di stabilirsi nella pianta dopo l'applicazione in campo come biofertilizzanti o agenti di biocontrollo.
La possibilità di passaggio da piante madri a piante figlie dei batteri a funzione PGP è stata testata da Guerrero-Molina et al., (2011) con i due ceppi PGPB di Azospirillum brasilense, REC3 e PEC5 su piante di fragola (Fragraria x ananassa Duch). Entrambi i ceppi sono stati in grado di migliorare la crescita e la resa delle colture. I ceppi REC3 e PEC5 non solo colonizzano in modo efficace la rizosfera, esplicando le loro funzioni, ma sono anche in grado di passare dalle piante madri ai nuovi impianti figli, per mezzo degli stoloni. Ciò garantisce la possibilità di avere numerose generazioni di piante in vivaio già inoculate, contribuendo così ad una pratica agricola più sostenibile
Il lavoro di Banchio et al. (2008) ha evidenziato invece gli effetti della colonizzazione dei ceppi Pseudomonas fluorescens WCS417r, Bradvrhizobium sp, B. subtilis 09 e Sinorhizobium meliloti Rm1021 su piante di Origanum majorana L.. Si è evidenziato un incremento della biomassa ed un miglioramento della composizione qualitativa e quantitativa degli oli essenziali. In particolare, rispetto al controllo senza inoculi batterici, le piante bioinoculate con P. fluorescens presentavano una percentuale significatamene più elevata di olio essenziale. Questo risultato evidenzia il potenziale impiego commerciale di questo batterio per la coltivazione di O. majorana. Sempre nella specie Origanum la biofertilizzazione con Azospirillum brasiliense ha mostrato un significativo aumento della concentrazione di terpeni (Banchio et al., 2010).
23 L’azione biopesticida dei PGPB, esplicata dalla produzione in terpeni, è stata testata anche in O. con i ceppi Pseudomonas monteilii CRC1, Cronobacter dublinensis CRC3 e Bacillus spp. Il significativo incremento dei livelli di acido rosmarinico, determinato dall’impiego del bioinoculo, ha mostrato di inibire la proliferazione di Pseudomonas aeruginosa, un batterio del suolo opportunista e patogeno per l’uomo (Pal Bais et al., 2002).
L’acido rosmarinico ha mostrato attività antimicrobica contro una serie di microrganismi terricoli ma, particolari effetti deleteri sono stati riscontrati nei confronti di Pseudomonas aeruginosa, suggerendo l’uso di questo bioinoculo per migliorare il rendimento e la gestione della malattia per i coltivatori biologici di basilico (Singh et al., 2013). L’incremento del peso fresco e l’accumulo in oli essenziali in O. basilicum è stato inoltre riscontrato nella produzione di cultivar bioinoculate con PGPB (Banchio et al., 2009).
Sebbene sia stato evidenziato che alcuni PGPB sono in grado di interagire autonomamente con le radici delle piante, i dati relativi all'intensità di popolazione, ne rilevano una buon funzionamento se in associazione alle ife degli AMF (Hartmann et al., 2009). Infatti, la produzione di polisaccaridi extracellulari, da parte di ceppi di Azospirillum e Rhizobium ad attività PGPB, ha mostrato di migliorare, in modo significativo, l'adesione di questi batteri alle strutture fungine e quindi alle radici. Questa adesione può influenzare in modo significativo il movimento dei PGPB verso nuove rizosfere (Bianciotto et al., 2001b).
Kim et al. (2010) hanno evidenziato che gli effetti sinergici tra Methylobacterium oryzae e diverse specie di funghi AM (Acaulospora longula, Glomus clarum e Glomus. intraradices) influenzano la crescita delle piante e il contenuto di clorofilla. Inoltre, il tasso di colonizzazione delle radici da parte dei funghi AM, il numero di spore e l’assorbimento della pianta in macro- e micro-nutrienti sono stati incrementati mediante l'inoculazione combinata dei due inoculi fungino e batterico.
Le interazioni tra PGPR e AMF, tuttavia, non sono sempre positive. Infatti, è stata riportata la possibilità di interazioni ininfluenti o negative, a causa della variabilità nella compatibilità funzionale tra le componenti microbiche (Bhromsiri et al., 2010).
24 La forza del legame batterico alle ife degli AMF è differente nelle diverse fasi fisiologiche di sviluppo dei funghi. Si assiste, infatti, a un attacco elettrostatico debole nella prima fase, seguito da un successivo forte attaccamento, dipendente dalla produzione di cellulosa o altri prodotti dai batteri extracellulari (Artursson et al., 2006). Questa ipotesi è supportata dalla minore adesione dei mutanti batterici che, in presenza di ife AM, non sono in grado di produrre tali composti organici, (Artursson et al., 2006).
Poiché alcuni ceppi batterici come Pseudomonas spp. sono in grado di colonizzare sia le radici delle piante che le ife AM, è stato suggerito che i processi relativi possono essere relativamente simili.
I PGPB e i funghi AM possono anche influenzare la decomposizione della materia organica nel suolo, che risente della loro produzione in enzimi (de Boer et al,. 2005). Esistono alcuni batteri (fosforo solubilizzanti) che sono in grado di migliorare l’assorbimento del fosforo in alcune specie di AMF (Zabihi et al., 2010; Salimpour et al., 2010). Ad esempio il fungo G. intraradices e i batteri dal genere Paenibacillus (P. macerans e P. polymyxa) sono stati esaminati in un esperimento in serra con Cucumis sativus. La presenza di G. intraradices è stata associata ad un aumento dei livelli di deidrogenasi e alla maggiore disponibilità di fosforo nel substrato, suggerendo che la formazione di micorrize accelera la decomposizione della materia organica con conseguente mobilitazione di fosforo
(John Larsen et al., 2009).
Poiché la coltivazione di basilico è molto esigente in fertilizzanti, in particolare azoto e fosforo, i coltivatori sono portati ad applicare grandi quantità di fertilizzanti per ottenere rese elevate. In questo contesto l'utilizzo di biofertilizzanti, quali PGPB può migliorare la crescita delle piante di basilico e, l’interazione con gli AMF, può incrementare la resa e la qualità produttiva, grazie ad un maggiore assorbimento in fosforo (Sharafzadeh et al., 2011).
Una sperimentazione condotta in serra è stata eseguita per studiare gli effetti dell’inoculazione con Glomus fasciculatum e i due batteri Bacillus megaterium e P. fluorescens con caratteri PGP, sulla crescita e biomassa di basilico coltivato in serra. È stato rilevato che l’inoculo singolo e doppio aumentano la crescita e la biomassa della specie, rispetto alle piante non inoculate. Il miglior risultato è stato
25 evidenziato dal consorzio di tutti e tre gli organismi, per quanto riguarda la valorizzazione in altezza della pianta, il numero di rami, la resa e il contenuto tissutale di fosforo, la percentuale di colonizzazione micorrizica e il numero di spore nel terreno (Hemavathi et al., 2006).
Sayed et al. (2011) hanno studiato l'effetto dell’inoculazione con B. subtilis e/o AMF su tre cultivar di basilico dolce (cultivar locale, Nano Compatt e Red Bordaux). I parametri esaminati sono stati la crescita della pianta, la percentuale in olio essenziale e l'assorbimento dei nutrienti a diversi livelli di stress salino. I risultati hanno evidenziato che l'elevato livello di salinità causa la riduzione in altezza delle piante, dei pesi freschi e secchi della pianta, del numero di rami, della percentuale in olio essenziale e del contenuto in N, P e K nelle foglie in tutte e tre le cultivar di basilico. L’inoculazione con B. subtilis e/o i funghi micorrizici ha mostrato effetti positivi sulla crescita, sulla percentuale in olio e sull'assorbimento dei nutrienti. La colonizzazione micorrizica ha manifestato effetti, in genere, più pronunciati rispetto a B. subtilis; mentre la doppia inoculazione con B. subtilis e AMF ha fornito maggiore tolleranza alla salinità. Questo lavoro ha indicato che l'inoculazione di cultivar di basilico con B. subtilis e AMF può indurre un incremento della tolleranza del basilico alla salinità.
1.5 Scopo della tesi
Le piante aromatiche hanno da tempo dimostrato di possedere spiccate attività antiossidanti, suggerendo, quindi, potenziali benefici per la salute umana. Tali attività sono ascrivibili a composti noti come fitochimici che includono polifenoli e ascorbato, di grande interesse per il loro valore nutraceutico. Il basilico (Ocimum basilicum L.) è una specie orticola introdotta in diverse aree del Mediterraneo e, sebbene considerata una coltura di nicchia, viene coltivata su una vasta superficie, per scopi sia alimentari che farmaceutici. Come la maggior parte delle piante alimentari, il basilico è in grado di stabilire simbiosi mutualistica con microrganismi della rizosfera, i funghi micorrizici arbuscolari (AMF), che svolgono un ruolo chiave per la nutrizione e la salute delle piante. L’attività dei
26 funghi micorrizici è positivamente influenzata dalle comunità batteriche della micorrizosfera, che vivono strettamente associate alle spore e alle ife degli AMF. Lo scopo della presente tesi è stato quello di saggiare l’impiego di associazioni di AMF e batteri ad essi associati come biofertilizzanti e biostimolanti. A tale scopo, nell’ambito di una prova in vaso con la pianta O. basilicum cv Tigullio, è stato utilizzato il fungo R. intraradices IMA6 inoculato da solo e/o in combinazione con 5 diverse specie batteriche precedentemente isolate dalle spore del fungo e selezionate per le loro spiccate attività PGP. Sono state effettuate analisi microbiologiche per la determinazione della colonizzazione micorrizica e analisi chimiche e biochimiche per la valutazione del contenuto in azoto e fosforo, della capacità antiossidante (mediante determinazione di fenoli e acido ascorbico) nonché dello stato fisiologico della pianta (mediante valutazione della fluorescenza della clorofilla a). L’effetto del bio-inoculo sulla diversità della comunità batterica della micorrizosfera è stato inoltre analizzato mediante l’utilizzo di tecniche molecolari coltura-indipendenti (PCR-DGGE).
27
2. MATERIALI E METODI
2.1 Ceppi microbici
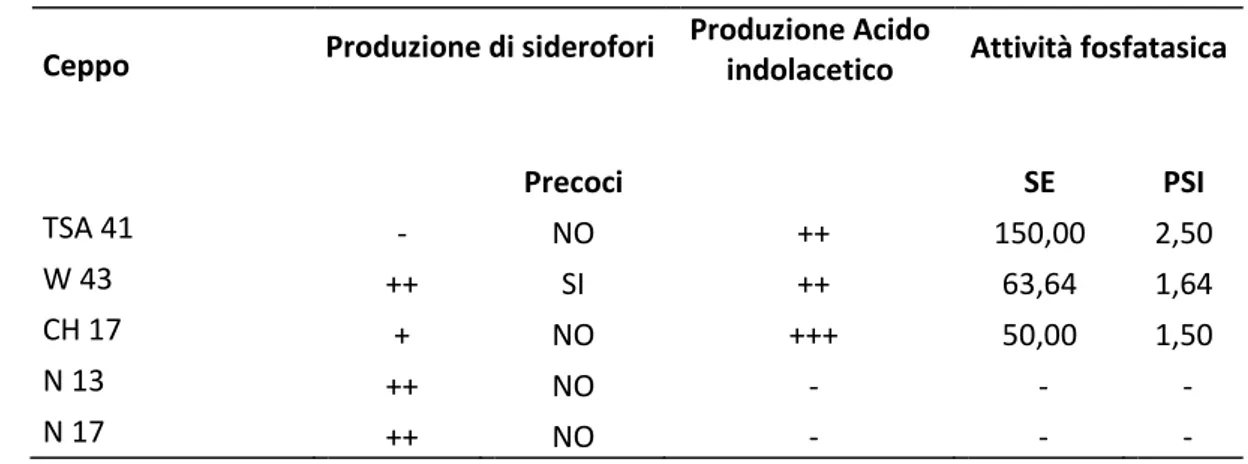
In un precedente lavoro di tesi (Bonari, 2014) sono stati isolati e analizzati per le loro caratteristiche PGP (Plant growth-promoting) ceppi batterici associati alle spore del fungo micorrizico arbuscolare R. intraradices IMA6. Questi sono stati inseriti nella collezione del Dipartimento di Scienze Agrarie, Alimentari e Agro-ambientali (laboratori di microbiologia) e mantenuti a -80°C in glicerolo al 20% . Tra questi, nell’ambito della presente tesi, sono stati selezioni 5 ceppi con spiccate caratteristiche PGP (Tabella 3) che sono stati identificati e utilizzati come inoculo per la prova in vaso.
Tabella 3: Ceppi microbici selezionati e loro caratteri PGP
Ceppo Produzione di siderofori
Produzione Acido
indolacetico Attività fosfatasica
Precoci SE PSI TSA 41 - NO ++ 150,00 2,50 W 43 ++ SI ++ 63,64 1,64 CH 17 + NO +++ 50,00 1,50 N 13 ++ NO - - - N 17 ++ NO - - -
I ceppi batterici sono stati rivitalizzati e mantenuti sui medesimi mezzi di coltura utilizzati per il loro isolamento.
2.1.1.Trypticsoy agar (TSA)
Il TSA è un mezzo generico utilizzato per la crescita di batteri aerobi coltivabili, impiegato anche per il rilevamento di Bacillus spp., la cui composizione è riportata in Tabella 4.
28
Tabella 4: Composizione TSA (Trypticsoy agar)
TSA Tryptic soy agar (Fluka)
Tryptic soy broth 30 g L-1
Bacteriological agar 20 g L-1
A seguito della sterilizzazione, 121°C per 15 minuti, sono stati addizionati gli antibiotici riportati in Tabella 5.
Tabella 5: Antibiotici TSA (Trypticsoy agar)
Cicloeximide 10 mL L-1
Nistatina 5 mL L-1
2.1.2. Mezzo minimo con chitina colloidale
Il mezzo minimo con chitina è stato utilizzato per la coltura dei batteri chitinolitici, i quali usano la chitina come unica fonte di carbonio (Souza et al., 2009); la sua composizione, per litro d’acqua milliQ, è riportata in Tabella 6.
Tabella 6: Composizione mezzo con chitina colloidale
Mezzo minimo con chitina colloidale
Chitina colloidale 0,3% (NH4)2SO4 1 g L-1 KH2PO4 0,2 g L-1 K2HPO4 1,6 g L-1 MgSO4- 7H2O 0,2 g L-1 NaCl 0,1 g L-1 FeSO4 7 H2O 0,01 g L-1 CaCl2 0,02 g L-1
29 Il mezzo è stato sterilizzato a 121°C per 15 minuti e addizionato degli antibiotici di Tabella 5.
2.1.3. Waksman agar
Il Waksman agar è un mezzo selettivo, usato per il rilevamento degli attinobatteri. Questo terreno colturale ha la composizione riportata in Tabella 7.
Tabella 7: Composizione Waksman agar
Waksman agar
Glucosio 10,0 g L-1
Cloruro di sodio 5,0 g L-1
Bacteriological Peptone (Oxoid) 5,0 g L-1
Estratto di carne Lab-Lemco (Difco) 3,0 g L-1
Agar bacteriological N.3 (Oxoid) 20 g L-1
A seguito dell’aggiunta di tutti i componenti, mostrati in Tabella 5, all’acqua deionizzata, il mezzo è stato portato a pH a 7.0 e messo a sterilizzare in autoclave a 121° C per 20 minuti. Al termine della sterilizzazione il mezzo è stato addizionato degli antibiotici per litro, riportati in Tabella 8:
Tabella 8: Antibiotici Waksman
Cicloeximide 10 mL L-1
Nistatina 5 mL L-1
Polimixina 5 mg L-1
2.1.4. Nitrogen free mineral salts
I microrganismi azoto fissatori sono stati fatti crescere su mezzo selettivo Nitrogen-free mineralsalt (N-free), la cui composizione è riportata in Tabella 9.
30
Tabella 9: Composizione Nitrogen-freemineralsalt (N-free) Nitrogen- free mineral salts
Soluzione stock salina 5 mL L-1
CaCO3 5,0 mg L-1
Agar Nobile N.1 20 g L-1
Per preparare il mezzo Nitrogen- free mineralsalts è stata utilizzata acqua milliQ. Inoltre, anche la soluzione salina di cui è composto, ha la composizione di Tabella 10 per litro di acqua milliQ:
Tabella 10: Soluzione Stock Salina
Soluzione salina MgSO4 7H2O 25 g L-1 KH2PO4 50 g L-1 NaCl 25 g L-1 FeSO4 7H2O 1 gL-1 MnSO4 4H2O 1 g L-1
La soluzione di Tabella 10 è stata sterilizzata per filtrazione e mantenuta a temperatura ambiente.
A seguito della messa a punto del pH, fino al valore di 7.2, il mezzo è stato sterilizzato in autoclave a 121°C per 15 minuti. Dopo la sterilizzazione è stata addizionata la soluzione zuccherina 100 mL L-1 e i supplementi per litro di mezzo riportati in Tabella 11.
Tabella 11: Antibiotici N-free (Nitrogen-freemineralsalt)
Cicloeximide 10 mL L-1
Nistatina 5 mL L-1
Per l’identificazione dei ceppi batterici sono stati utilizzati metodi molecolari quali estrazione del DNA (White et al., 1990), amplificazione del 16S rDNA (Lane 1991; Weisburg et al., 1991) e sequenziamento dell’amplificato 16S rDNA.
31
2.2.1 Estrazione del DNA
Per estrarre il DNA batterico è stato usato il kit “MasterPureTM
Yeast DNA Purification” dell'Epicentre.
Un volume di 1 mL delle colture cresciute in mezzo liquido è stato prelevato, trasferito in tubi eppendorf e centrifugato a 10000 rpm per 10 minuti. Il pellet è stato risospeso in 300 µL di Yeast Cell Lysis Solution ed incubato a 65°C per 15 minuti e successivamente in ghiaccio per 5 minuti. Quindi sono stati aggiunti 150 µL di MPC Protein Precipitation Reagent e la sospensione è stata agitata tramite vortex per 10 secondi e successivamente centrifugata a 10000 rpm per 10 minuti. Il surnatante è stato prelevato e trasferito in un tubo Eppendorf al quale è stato aggiunto un ugual volume di isopropanolo (Sigma-Aldrich) e incubato a temperatura ambiente per 5 minuti. Il campione è stato nuovamente centrifugato a 10000 rpm per 10 minuti. Eliminato il surnatante il pellet così ottenuto è stato lavato con etanolo al 70% e centrifugato a 10000 rpm per 5 minuti.
Dopo aver eliminato il surnatante, il pellet è stato lasciato ad asciugare all’aria e risospeso in 50 µL di TE e mantenuto in frigo a 4°C.
La reazione di amplificazione della regione 16S è stata condotta utilizzando lo strumento iCycleriQMultiocolor Real-Time PCR Detection System (BIORAD). La reazione di amplificazione è stata effettuata in un volume finale di 25 µL, contenente 1 µL di stampo genomico (1:10), 0,2 mM di dnTPs (Finnzymes), 2,5 µL di 10X Finnzymes (1X buffer: 50 mM di KCl, 10 mM di Tris-HCl pH 8.8 25°C, 1,5 Mm MgCl2, 0,1% di Triton X-100), 1.25 U di Taq DNA polimerasi
(Finnzymes), 0,2 µM di ciascuno dei primers 27f (5’-GAG AGT TTG ATC CTG GCT CAG-3’) e 1495r (5’-CTA CGG CTA CCT TGT TAC GA-3’).
32
Tabella 12 Programma di amplificazione
Denaturazione Amplificazione (35 cicli) Estensione finale Temperatura 95°C 94°C 54°C 72°C 72°C Tempo 2’ 1’20’’ 1’ 1’30’’ 5’
La presenza degli amplificati è stata confermata mediante elettroforesi in tampone TBE 1X su un gel di agarosio alla concentrazione del 2% contenente 0,5 µg mL-1 EtBr. A 5 µL di amplificato sono stati aggiunti 3 µL di tampone di caricamento e utilizzato un marker di riferimento con frammenti a peso molecolare noto 100 pb DNA Ladder (BioLabs, New England). Le soluzioni, in acqua distillata, utilizzate per le corse elettroforetiche su gel di agarosio, sono riportate in Tabella 13 e 14.
Tabella 13: Tampone Tris-borato
Trizma 54 g L-1
Acido borico 27,5 g L-1
EDTA 0,5M(pH8) 20 mL L-1
Tabella 14:Gel LoadingDye, Blue (1X)
Blu di bromofenolo (1X) 2,5 g L-1
Ficoll (tipo 400) (1X) 150 g L-1
Gli amplificati sono stati acquisiti mediante il programma LISCAP per IMAGE-MASTER VDS system, e analizzati con il programma Image Master Elite (Pharmacia Biotech).
2.2.2 Sequenziamento del 16S rDNA e analisi delle sequenze
Gli amplificati purificati dai dNTPs e dai primers in eccesso, utilizzando le colonnine “EuroGOLDCycle-Pure Kit” (Euroclone), sono stati sequenziati presso BMR Genomics (Padova, Italia). Le sequenze sono state analizzate per
33 determinare l’omologia con le sequenze depositate in banca dati utilizzando il programma BLAST (http://www.NCBI.nim.nih.gov/). Le sequenze sono state allineate usando Muscle (Edgar, 2004a; 2004b) e gli alberi filognetici sono stati costruiti usando il metodo Neighbor-Joining (modello di correzione Kimura) con il software Mega 5.2 (http://www.megasoftware.net/). I valori di bootstraps sono stati calcolati su 1000 replicati, per fornire le stime di fiducia a 1000 repliche bootstrap.
2.3. Prova in vaso
I ceppi batterici sono stati utilizzati separatamente e unitamente al fungo micorrizico R. intraradices IMA6 come bio-inoculi in prove in vaso con la specie vegetale Ocimum basilicum cv. Tigullio.
2.3.1 Produzione dell’inoculo micorrizico
Per la moltiplicazione dell’isolato fungino IMA6, sono stati allestiti 4 vasi da 8 L ciascuno, utilizzando come piante ospiti Sonchus asper, Helianthus annuus e Trifolium alexandrinum, specie altamente micotrofiche, in grado di stabilire simbiosi MA con l’endofita utilizzato. La coltura è stata condotta in serra con adeguate e periodiche annaffiature senza l’aggiunta di fertilizzanti. Per la produzione sono stati utilizzati 4 L di suolo agrario proveniente dal Centro sperimentale di ortofloricoltura, S. Piero a Grado, Pisa e 4 L di Terragreen (argilla calcinata, Oil Dri). Prima dell’utilizzo il terreno è stato setacciato manualmente utilizzando una griglia con fori dal diametro di 0,5 centimetri. Successivamente, entrambi i materiali sono stati sottoposti a due cicli di sterilizzazione ciascuno a 121°C per 30 minuti. Come inoculo starter è stato usato materiale proveniente dalla collezione del DiSAAA-a, contenente spore, radici micorrizate e micelio dei ceppi fungini prescelti. Il substrato è stato realizzato miscelando suolo agrario, Terragreen e lo starter in rapporto 1:1:2 v:v:v. Dopo 4 mesi, eliminata la parte aerea delle piante, il substrato è stato rimescolato e omogeneizzato con le radici tagliate in frammenti di circa 1 cm.