La masticazione umana è un fenomeno molto complesso, nel cui funzionamento entrano in gioco numerosi fattori. Oltre alla funzione principale di triturazione dei cibi e inizio della digestione, il sistema di masticazione umana, costituito dall’apparato stomatognatico, è interessato in vari aspetti funzionali dell’organismo, come la deglutizione, la fonazione e la postura. Conseguenza di questa complessità è la difficoltà a costruire una replica artificiale profondamente fedele di questo sistema, in quanto sarebbe necessaria l’integrazione con altri elementi che simulino le funzioni cui la masticazione umana è associata.
In questo capitolo vengono illustrati brevemente gli aspetti dell’apparato stomatognatico che più ci interessano: i movimenti della mandibola e il sistema neurosensoriale di controllo. Questo può essere utile per avere un quadro, sebbene non esaustivo, del funzionamento del meccanismo biologico cui ci si vuole ispirare, e per capire quanto le inevitabili semplificazioni del sistema artificiale incidano sulla sua fedeltà di simulazione.
1.2 Movimenti della mandibola umana
Nell’uomo i movimenti della mandibola rispetto alla mascella sono realizzati tramite le articolazioni temporo-mandibolari. Si tratta di condiloartrosi nelle quali il movimento avviene attraverso la rotazione e traslazione dei capi articolari della
mandibola, detti condili mandibolari, all’interno della fossa mandibolare dell’osso temporale, cavità ossea delimitata posteriormente dalla parte timpanica dell’osso temporale e anteriormente dalla radice del processo zigomatico. La forma della testa dei condili e della fossa sono discordanti, ma l’articolazione è resa coerente dal disco articolare, struttura di natura fibrosa posta tra i due capi dell’articolazione e aderente ad entrambi [5].
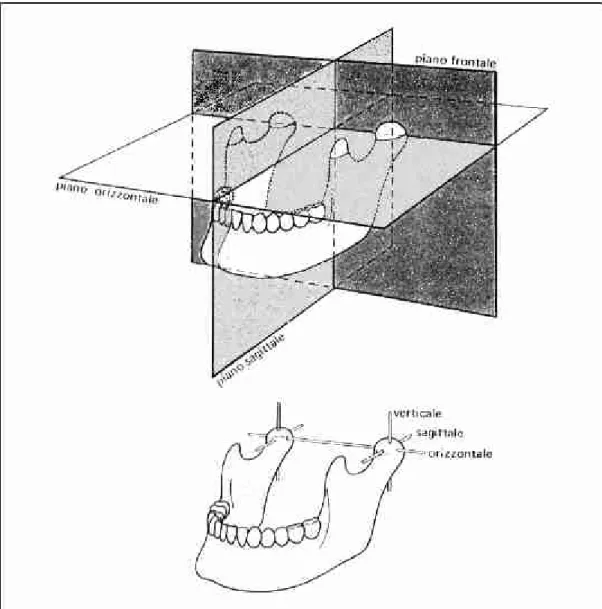
Figura 1.1: piani di riferimento per i movimenti della mandibola e assi di rotazione dei condili.
riferimento e gli assi attorno ai quali avvengono le rotazioni dei condili (figura 1.1). I piani utilizzati, tra loro ortogonali, sono:
• il piano frontale, che è disposto in senso verticale e divide il cranio in una parte anteriore e una posteriore;
• il piano sagittale mediano, posto anch’esso in senso verticale, che divide il cranio in una parte destra e una sinistra;
• il piano orizzontale, che separa la parte inferiore da quella superiore del cranio. Le rotazioni dei condili possono avvenire intorno agli assi verticale, antero-posteriore o sagittale e orizzontale; quest’ultimo è comune ai due condili, gli altri due sono entrambi monocondilari.
A questo punto possiamo descrivere brevemente i tre movimenti elementari della mandibola [6]:
• Moto di apertura e chiusura (o di abbassamento e innalzamento). È un movimento che avviene nel piano sagittale tramite una rotazione consensuale dei condili attorno all’asse orizzontale e una loro traslazione. Il moto di apertura può cominciare dalla posizione di massimo contatto delle superfici occlusali dei denti delle due arcate, detto occlusione centrica, oppure dalla posizione di riposo, in cui la mandibola è leggermente aperta; il passaggio dalla posizione di occlusione centrica a quella di riposo avviene tramite una rotazione dei condili attorno all’asse orizzontale di circa 2°. Il moto di apertura comincia con una rotazione dei condili dall’avanti all’indietro (con una contemporanea traslazione minima), seguita da una traslazione verso avanti e verso il basso e, infine, da un’ulteriore rotazione che può portare la mandibola in posizione di massima apertura, nella quale gli incisivi antagonisti normalmente distano tra loro 5-6 cm. Partendo da questa posizione, il moto di chiusura comincia con una rotazione in senso inverso, seguita da un movimento combinato di rotazione e traslazione in direzione posteriore e verso l’alto dei condili.
• Moto di protrusione e retrusione. È un movimento in cui la mandibola scorre nel piano orizzontale in direzione antero-posteriore tramite una rotazione e una minima traslazione dei condili, che anche in questo caso compiono lo stesso tipo di
spostamento. Nella protrusione la testa del condilo e il disco articolare si portano avanti e in basso, nella retrusione avviene lo spostamento opposto.
• Moto di traslazione laterale. Questo movimento consente lo spostamento della mandibola verso destra e verso sinistra sul piano orizzontale. Stavolta i condili sono soggetti a due spostamenti diversi: uno, detto condilo ruotante, fa praticamente da perno, compiendo una rotazione con movimento laterale e posteriore intorno all’asse verticale, mentre l’altro, detto condilo orbitante, si sposta anteriormente e verso il basso descrivendo un piccolo arco di cerchio. Ad esempio, nel caso di un movimento verso destra, il condilo orbitante è quello sinistro. Lo spostamento laterale della mandibola è anche detto adduzione, il ritorno alla posizione di riposo abduzione.
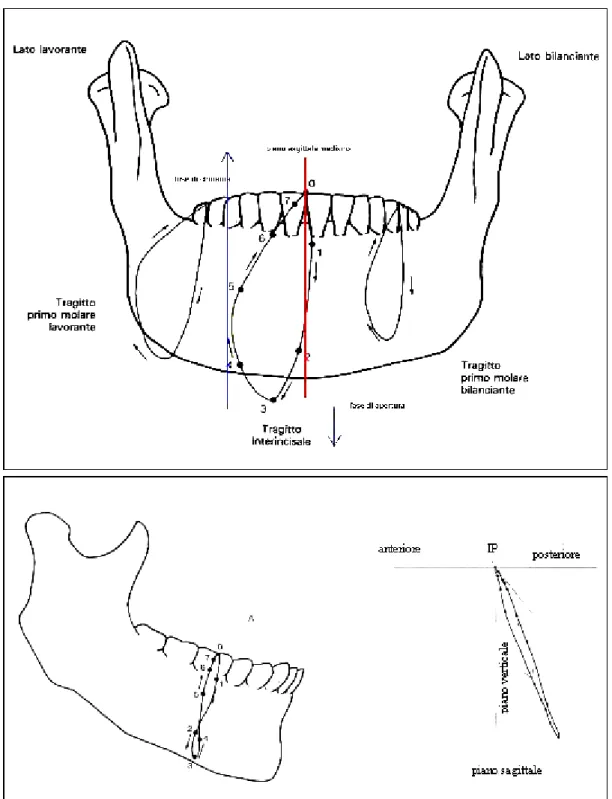
I movimenti compiuti dalla mandibola durante la masticazione, pur variando notevolmente da individuo a individuo, rispondono ad un modello caratteristico coordinato e scomponibile nei tre piani di riferimento. In particolare, sul piano orizzontale si hanno spostamenti laterali; sul piano sagittale la mandibola compie dei movimenti di innalzamento e abbassamento; infine, sul piano frontale si ha una combinazione dei due movimenti precedenti, che dà origine ad una traiettoria più o meno circolare. Quest’ultima è asimmetrica, a causa del fatto che la masticazione avviene tipicamente solo da un lato, detto di lavoro, mentre l’altro è detto di bilanciamento. In figura 1.2 sono rappresentate le traiettorie di alcuni punti della mandibola durante la masticazione, sui piani frontale e sagittale.
Tutti questi movimenti, combinati tra loro, danno origine al ciclo masticatorio, che può essere suddiviso in tre fasi [6]:
• fase di apertura, in cui la mandibola si abbassa, prima lentamente poi più velocemente, e il cibo viene inserito nella cavità orale;
• fase di chiusura, divisa anch’essa in rapida e lenta, in cui la mandibola si solleva e ruota in direzione del lato di lavoro;
• fase occlusale, che occupa la maggior parte del ciclo e corrisponde al periodo di lavoro in cui si esercita pressione sul cibo interposto tra le due arcate. In quest’ultima fase avviene anche il contatto tra le superfici occlusali dei denti antagonisti, e la
mandibola si inclina dapprima facendo perno sul bolo, poi per il contatto tra i denti del lato di bilanciamento.
Figura 1.2: traiettorie dei denti durante un ciclo di masticazione. Sopra: traiettorie degli incisivi, del primo molare lavorante e del primo molare bilanciante sul piano frontale. Sotto: schema di un movimento di masticazione sul piano sagittale.
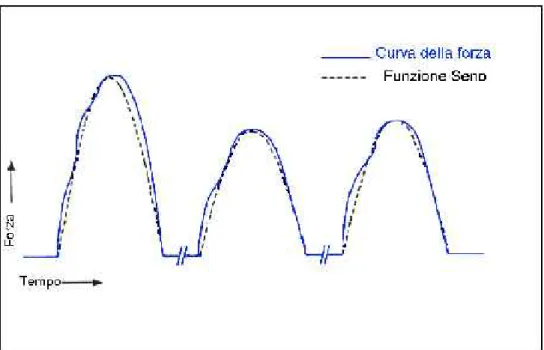
Per quanto riguarda le forze agenti sui denti, possono raggiungere valori notevoli nella chiusura delle arcate: fino a 25 kg in corrispondenza degli incisivi e addirittura fino a 80 kg in corrispondenza dei molari [6]. L’andamento di tali forze durante un ciclo di masticazione può essere approssimato con la semionda positiva di una funzione sinusoidale, come rappresentato nella figura seguente. La frequenza di tale ciclo è compresa tra 0.5 e 5 Hz.
Figura 1.3: profilo delle forze agenti sui denti dell’arcata mandibolare durante un ciclo di masticazione (linea continua), con sovrapposta la semionda positiva di una sinusoide (linea tratteggiata).
1.2 Il controllo nervoso della masticazione
Le modalità in cui il sistema nervoso controlla la masticazione non sono ancora completamente chiare. Tuttavia, si conoscono vari importanti meccanismi cui è utile accennare, sia per capire quanto i dispositivi di traduzione utilizzati nel sistema artificiale siano riconducibili a qualche tipo di recettore biologico, che per trovare ispirazione per un controllo neuromorfo di tale sistema.
1.2.1 Controllo della muscolatura mandibolare e recettori della
masticazione
I muscoli interessati nella masticazione sono classificati in base al tipo di movimenti che consentono alla mandibola. Pertanto abbiamo: muscoli elevatori (massetere, temporale e pterigoideo interno) ed abbassatori (digastrico, pterigoideo esterno, miloioideo, genioioideo); muscoli protrusori (pterigoideo interno ed esterno) e retrusori (temporale e digastrico); muscoli abduttori e adduttori (pterigoideo interno ed esterno).
La sequenza di azionamento di questi muscoli durante la masticazione è molto complessa, e il numero di movimenti e l’ampiezza delle contrazioni variano notevolmente da un individuo all’altro.
I muscoli masticatori, come tutta la muscolatura scheletrica dell’organismo, sono comandati da motoneuroni, ognuno dei quali innerva varie fibre muscolari. In particolare, quelli implicati nel movimento dei muscoli masticatori sono di tipo α, gli α-motoneuroni.
In questi muscoli sono presenti vari tipi di recettori, tra i quali sono particolarmente importanti i fusi neuromuscolari, strutture diffuse in tutto l’organismo (specialmente nella muscolatura associata all’attività posturale) e presenti, per quanto riguarda il sistema mandibolare, solo nei muscoli elevatori [7].
Un fuso neuromuscolare, che misura tipicamente dai 3 ai 10 mm di lunghezza, è costituito da un certo numero (solitamente da 3 a 12, ma nei muscoli masticatori possono arrivare anche a 36, vedi [8]) di piccole fibre muscolari intrafusali, le cui estremità sono collegate alle fibre extrafusali circostanti. La parte recettrice del fuso si trova nella zona centrale, dove le fibre intrafusali sono sprovviste di elementi contrattili (i filamenti di actina e miosina), ma sono presenti le terminazioni dei nervi sensitivi. Queste vengono eccitate dallo stiramento della zona centrale del fuso, che può avvenire in seguito all’allungamento dell’intero muscolo o anche, se la lunghezza di quest’ultimo non si modifica, a una contrazione delle porzioni laterali delle fibre intrafusali. In ogni fuso sono presenti due tipi di terminazioni sensitive: quelle primarie (una per fuso), che sono fibre nervose di tipo Ia, le più veloci dell’intero organismo, e quelle secondarie,
più lente e con un diametro inferiore. Le terminazioni secondarie rispondono agli stimoli solo in modo statico, ovvero trasmettono segnali per tutto il tempo in cui il recettore rimane eccitato, mentre quelle primarie rispondono anche in modo dinamico, ovvero molto più intensamente in corrispondenza dell’aumento della lunghezza del fuso. Le fibre extrafusali sono innervate dagli α-motoneuroni, quelle intrafusali da fibre nervose efferenti di tipo γ, i γ-motoneuroni.
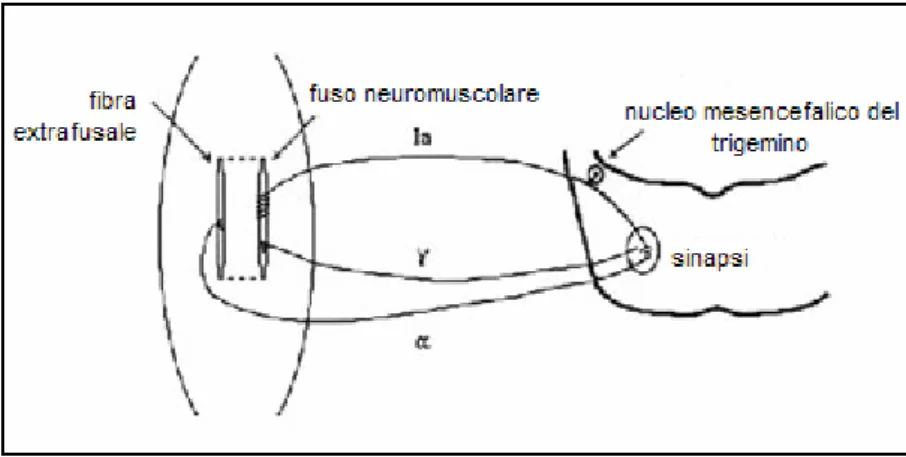
Gli α-motoneuroni sono collegati con la Ia afferente dello stesso muscolo in una modalità detta circuito monosinaptico di riflesso, in quanto i loro terminali distali formano un’unica sinapsi; a questa si aggiungono, in parallelo, le terminazioni delle fibre efferenti di tipo γ. La particolarità del sistema motorio mandibolare è che i corpi cellulari delle fibre afferenti non si trovano nel midollo spinale, come invece avviene per gli altri muscoli scheletrici, bensì nel Sistema Nervoso Centrale (SNC), in particolare nel nucleo mesencefalico del trigemino. Questa situazione è rappresentata nella figura seguente.
Figura 1.4: circuito monosinaptico di riflesso ed efferenti γ dei muscoli elevatori della mandibola.
Il sistema di funzionamento dei fusi neuromuscolari nei muscoli elevatori della mandibola implementa un controllo della lunghezza muscolare a feedback negativo: come detto sopra, la Ia afferente aumenta la sua attività in seguito allo stiramento della parte centrale del fuso, che può avvenire tramite l’allungamento dell’intero muscolo o la contrazione delle sezioni laterali del fuso dovuta all’eccitazione delle fibre γ; un
aumento dell’attività degli α-motoneuroni causa una contrazione dell’intero muscolo, ovvero una diminuzione dell’eccitazione della Ia afferente. Ad esempio, uno stiramento del muscolo attiva il fuso neuromuscolare che, a sua volta, attraverso il circuito monosinaptico tra motoneurone α e Ia afferente determina la contrazione del muscolo stesso, opponendosi al suo allungamento [9].
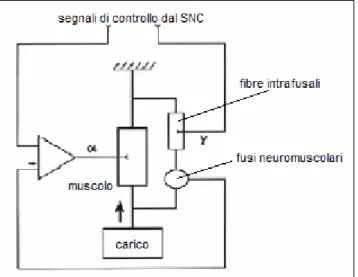
Si tratta di un meccanismo molto simile a quello utilizzato dall’organismo per il controllo della postura in cui, d’altronde, la masticazione è interessata [8]. Nel sistema mandibolare tale meccanismo serve, tra l’altro, per contrastare l’azione del peso della mandibola e di eventuali altre forze agenti su di essa, e mantenerla in posizione elevata. Lo schema di funzionamento può essere rappresentato come in figura 1.5. Le fibre di tipo γ servono, ad esempio, per compensare variazioni del peso agente sulla mandibola attraverso, in pratica, un movimento servo-assistito: se il carico mandibolare aumenta, si ha un allungamento del muscolo, che causa un aumento dell’attività dei motoneuroni α e, di conseguenza, una contrazione del muscolo che bilancia l’allungamento iniziale, consentendo al sistema di rimanere stabile.
Figura 1.5: schema di controllo del carico mandibolare tramite feedback negativo tra segnali del SNC e fusi neuromuscolari. Un aumento del carico mandibolare determina un allungamento del muscolo che, a sua volta, causa una maggiore attività dei motoneuroni α; questo provoca una contrazione del muscolo che bilancia l’allungamento iniziale.
Altri recettori interessati nella masticazione, con un ruolo importante soprattutto nella percezione cosciente del movimento e della posizione della mandibola, sono
situati nell’articolazione temporo-mandibolare. Dal punto di vista funzionale, questi recettori possono essere a rapido o a lento adattamento: i primi si attivano solo all’inizio di un movimento di elevazione o abbassamento della mandibola, i secondi rimangono eccitati per tutto il tempo in cui la mandibola non si trova nella situazione di riposo, anche se è ferma. Più in particolare, si tratta di organi di Ruffini e del Golgi, che fungono da meccanocettori statici, terminazioni di Vater-Pacini, che rappresentano quelli dinamici, e terminazioni nervose libere, che hanno varie funzioni, tra cui quella nocicettiva [8]. Questi e altri tipi di recettori sono diffusi anche nella pelle e nella mucosa facciale.
Un altro organo in cui vi sono varie tipologie di terminazioni sensitive è il legamento parodontale, ovvero il sistema di fibre che fissa il dente alla sua sede e che ammortizza le spinte pressorie verticali sul dente stesso. I recettori presenti in questo legamento, detti meccanocettori parodontali (PMR), rilevano intensità, direzione e velocità di applicazione delle forze che agiscono sul dente, oltre che stimoli termici, chimici e di dolore. I corpi cellulari dei PMR si trovano sul ganglio oppure sul nucleo mesencefalico del trigemino, in prossimità di quelli dei fusi muscolari dei muscoli elevatori; questo ha portato a supporre che ci sia un collegamento elettrico tra i corpi cellulari di questi due tipi di recettori. Inoltre, hanno entrambi delle proiezioni dirette sulla corteccia cerebellare, cosa che può essere spiegata dal fatto che per il controllo della postura mandibolare non sono sufficienti le informazioni provenienti dai fusi muscolari; queste, infatti, riguardano i movimenti ma non la posizione della mandibola, pertanto è necessaria l’integrazione tra tali segnali e quelli dei PMR, che indicano l’istante in cui avviene l’occlusione. In questo modo il cervelletto è in grado di regolare adeguatamente l’attività dei muscoli mandibolari, attraverso una calibrazione dei segnali nervosi sui fusi muscolari [8].
1.2.2 Riflessi del sistema mandibolare
Come altri apparati muscolo-scheletrici del corpo, anche quello mandibolare è soggetto a riflessi, ovvero a movimenti che non richiedono un’attivazione volontaria,
ma avvengono automaticamente al presentarsi di determinate condizioni. Esistono vari tipi di riflesso, i quali, proprio per essere considerati tali, devono avere delle proprietà in comune, ovvero per ognuno di essi si devono poter identificare: i recettori deputati alla rivelazione dello stimolo; il neurone primario afferente connesso al recettore; il suo corpo cellulare, che può trovarsi sia all’interno che all’esterno del SNC; i motoneuroni che controllano i muscoli interessati nel riflesso; le caratteristiche, in termini di tempistica, intensità e locazione, dello stimolo che causa il riflesso; la risposta a tale stimolo; la funzione del riflesso.
Dal punto di vista funzionale possiamo distinguere vari tipi di riflesso, due dei quali interessano la mandibola: il riflesso posturale o antigravità e quello protettivo o nocicettivo [9].
Il riflesso di chiusura della mandibola, ad esempio, è di tipo posturale: i recettori sono i fusi muscolari dei muscoli elevatori, i quali, in seguito all’apertura della mandibola, trasmettono un segnale la cui risposta consiste nella contrazione dei muscoli elevatori e la chiusura della mandibola. In questo modo riusciamo a tenere la mandibola in condizione di riposo nonostante l’azione della forza di gravità sulla mandibola stessa.
Un altro riflesso, stavolta di tipo protettivo, è quello di apertura della mandibola o linguomandibolare, che serve per proteggere i tessuti della cavità orale (ad esempio, è quello che causa l’apertura della bocca appena mordiamo inavvertitamente la lingua): in seguito a stimoli meccanici molto intensi o di dolore che colpiscono meccanocettori e nocicettori della cavità orale, in particolare i PMR, si ha il rilascio dei muscoli elevatori e l’apertura della mandibola.
Vi sono altri riflessi mandibolari, in particolare di protezione, per alcuni dei quali non sono ancora completamente chiare funzioni e struttura.
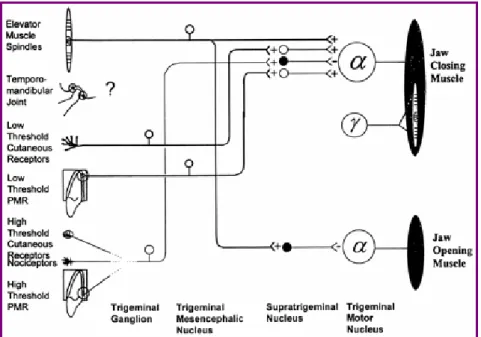
In figura 1.6 è rappresentato uno schema che riassume le connessioni nervose di questi riflessi.
1.2.3 Basi neurali del controllo del ciclo di masticazione
le caratteristiche dei riflessi mandibolari; non abbiamo ancora parlato, però, di come il ritmo di masticazione venga gestito dal SNC.
Nel 1917 Sherrington, studiando i riflessi di apertura e chiusura della mandibola nei gatti, avanzò l’ipotesi che tali riflessi costituissero il meccanismo di base della masticazione. Secondo la sua teoria, il ciclo masticatorio è dovuto a una catena di riflessi mandibolari: il processo viene attivato in seguito al posizionamento del cibo nella bocca, che provoca un riflesso di apertura in seguito al quale la mandibola si abbassa, causando a sua volta l’attivazione di un riflesso di chiusura che fa salire la mandibola fino all’occlusione delle arcate dentarie; la chiusura della bocca comprime il bolo alimentare contro le pareti della cavità orale, riattivando il riflesso di apertura della mandibola, e il ciclo si ripete.
Figura 1.6: diagramma di collegamento di muscoli, recettori e motoneuroni nel sistema mandibolare. A destra sono rappresentati i muscoli elevatori (in alto) e abbassatori (in basso); a sinistra i vari tipi di recettori; nella zona centrale i corpi cellulari dei motoneuroni e delle fibre sensitive [8].
Questa spiegazione è stata accettata per molti anni, nonostante contenesse varie contraddizioni abbastanza evidenti, prima fra tutte il fatto che la frequenza di masticazione è troppo bassa per poter essere generata da una catena di riflessi
mandibolari: infatti, i ritardi dei riflessi di apertura e chiusura della mandibola sono rispettivamente di 10 e 20 ms, perciò una loro combinazione darebbe origine ad una frequenza di masticazione di circa 33 Hz, mentre in realtà varia da 0.5 a 5 Hz. Inoltre, come abbiamo già visto, il ciclo masticatorio non consiste solamente in una sequenza ritmica di abbassamento ed elevazione della mandibola, ma è costituito da una complessa combinazione di movimenti, tra i quali anche quelli laterali e antero-posteriori [10].
Negli anni ‘70, grazie anche allo sviluppo di nuove tecniche di indagine, si è cominciato a far luce sul problema, arrivando alla conclusione che il ritmo masticatorio è generato da centri nervosi che svolgono la funzione di ”clock” interno e possono agire anche indipendentemente dalle afferenze periferiche, in modo molto simile a ciò che accade quando camminiamo o nuotiamo. Questi centri, che assumono complessivamente il nome di Central Pattern Generator (CPG), sono situati nella formazione reticolare midollare del tronco encefalico. Il CPG gestisce il ritmo della masticazione inducendo un’attivazione dei motoneuroni dei muscoli abbassatori e contemporaneamente un’inibizione di quelli dei muscoli elevatori durante la fase di apertura della mandibola; alternativamente, nella fase di chiusura inibisce i muscoli abbassatori ed eccita quelli elevatori.
Se lo schema di base del ritmo masticatorio è gestito dal CPG, i recettori periferici svolgono un ruolo fondamentale nella sua attivazione e nel modulare l’intensità, la frequenza, il tipo e la durata della masticazione in base alle caratteristiche del cibo, rendendo il ciclo adattabile alla situazione contingente. L’interazione tra CPG e recettori periferici può avvenire attraverso feedback sia positivi, ad esempio quando è necessario aumentare la forza di chiusura della mandibola per riuscire a masticare del cibo duro, che negativi, come quando si trova un sassolino tra i denti o ci si morde la lingua [10].
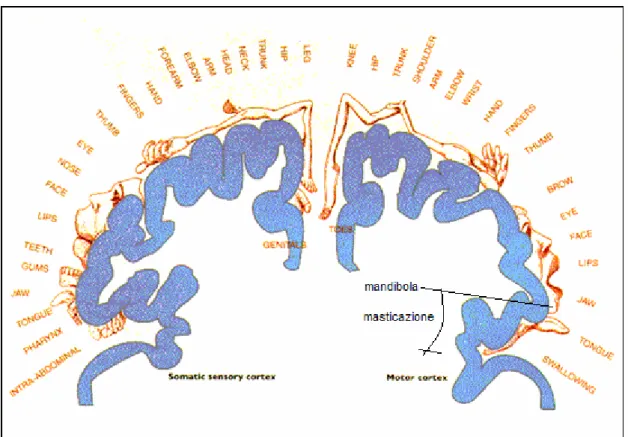
Un altro aspetto, anch’esso fondamentale, nel controllo della masticazione, è il ruolo della corteccia cerebrale, che consente l’attivazione volontaria dei muscoli masticatori. La corteccia cerebrale può fornire input ai muscoli mandibolari in modo sia diretto che indiretto. In particolare, la zona dedicata alla masticazione si trova nella corteccia
motoria primaria (figura 1.7) e, analogamente ad altri movimenti del corpo umano, il controllo corticale della masticazione consiste nell’attivazione dello schema funzionale memorizzato nel CPG. Vi sono anche delle fibre nervose efferenti ai muscoli mandibolari che provengono direttamente dalla corteccia cerebrale. Il controllo cerebrale viene attivato quando si decide volontariamente di iniziare o concludere l’atto masticatorio, oppure quando muoviamo la mandibola in un task cosciente avente un determinato scopo, come quando afferriamo un oggetto con i denti.
Un ruolo particolarmente rilevante nel fornire i segnali di feedback per la modulazione della masticazione è svolto dai PMR, che sono sensibili all’entità del carico agente sui denti [8].
È stato dimostrato, infine, che la masticazione può essere attivata anche in seguito alla stimolazione di altre zone cerebrali, come l’ipotalamo, l’amigdala e alcune zone della corteccia vicino alle aree sensoriali del gusto e dell’olfatto [11].
Figura 1.7: rappresentazione delle diverse zone del corpo nella corteccia somestesica (a sinistra) e motoria (a destra). In quest’ultima è indicata la zona deputata al controllo dei muscoli mandibolari